Primera semana del
desarrollo humano
(días 1 a 7)

FECUNDACIÓN

Ocurre en la ampolla de la trompa de Falopio y tiene tres fases
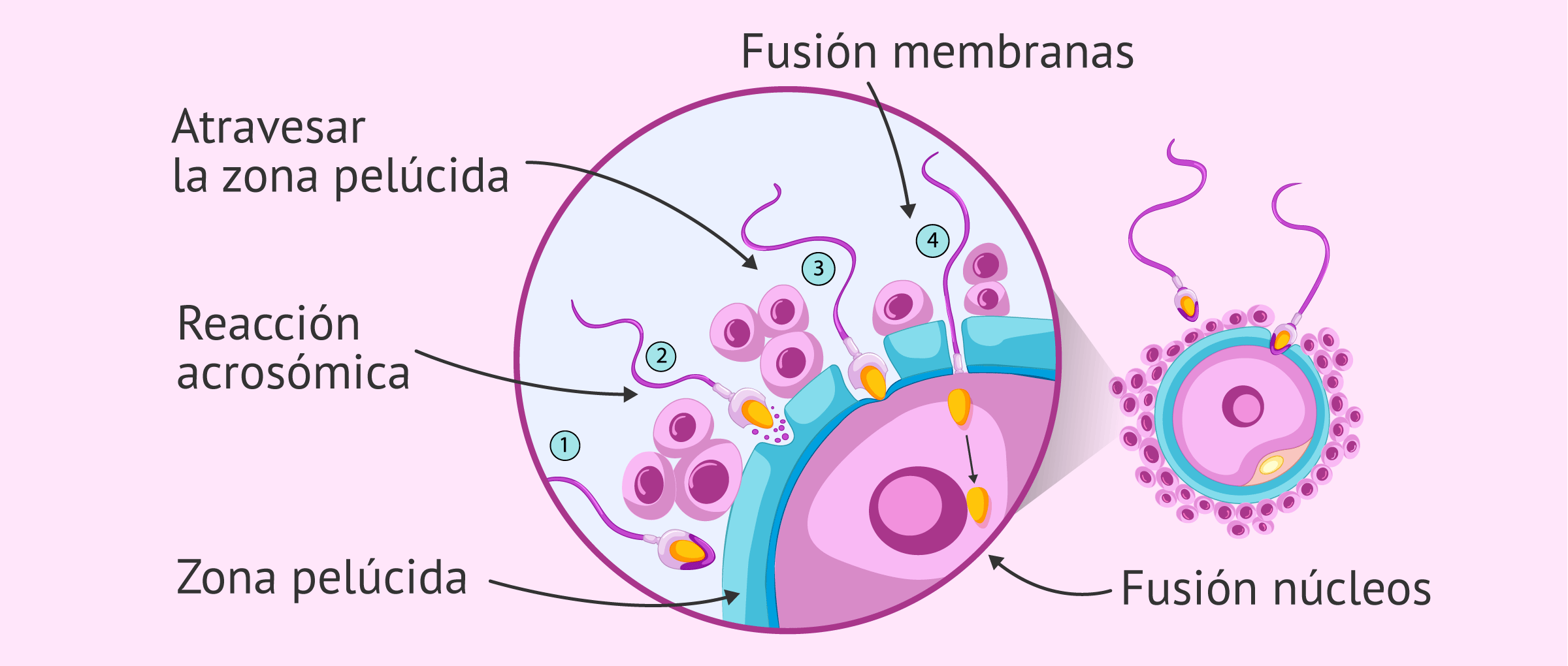
Fase 1: Penetración del espermatozoide en la corona radiada. La acción de las enzimas del espermatozoide y de la mucosa de la trompa de Falopio facilita este proceso.

Fase 2: Unión y penetración del espermatozoide en la zona pelúcida

La unión del espermatozoide ocurre por medio de la interacción entre las glucosiltransferasas y los receptores ZP3 situados en la zona pelúcida. Esta unión desencadena la reacción acrosómica, que consiste en la fusión entre la membrana acrosómica externa y la membrana plasmática del espermatozoide, que da lugar a la liberación de las enzimas acrosómicas
Las enzimas acrosómicas, en especial la acrosina, facilitan la penetración en la zona pelúcida. El contacto del espermatozoide con la membrana plasmática de un ovocito secundario desencadena la reacción cortical, que consiste en la liberación de gránulos corticales (lisosomas) desde el citoplasma del ovocito. Esta reacción cambia el potencial de la membrana plasmática del ovocito secundario e inactiva los receptores de los espermatozoides en la zona pelúcida. Estos cambios conforman el bloqueo poliespermático, el cuál deja impermeable la membrana plasmática del ovocito secundario a otros espermatozoides. De cualquier manera, la eficiencia del bloqueo poliespermático es cuestionable ya que la triploidía diándrica (un embrión con tres grupos de cromosomas, dos de los cuáles provienen del padre) es común.
Fase 3: Fusión de las membranas plasmáticas del espermatozoide y del ovocito, ocurre con la consiguiente ruptura de ambas membranas en la zona de fusión.
El espermatozoide entero (excepto la membrana plasmática) penetra en el citoplasma del ovocito
secundario detenido en la metafase de la meiosis II. El contenido nuclear del espermatozoide y el par de centriolos persisten, pero las mitocondrias y la cola del espermatozoide degeneran. El núcleo del espermatozoide se denomina pronúcleo masculino. Todas las mitocondrias del espermatozoide degeneran, y por ello todas las mitocondrias que se encuentran en el cigoto son de origen maternal (es decir, todo el DNA mitocondrial es de origen materno). El ovocito pierde su par de centriolos durante la meiosis por lo que la formación de un cigoto funcional depende del
par de centriolos del espermatozoide (una característica cardinal de la embriogénesis humana) para producir un centro organizador de microtúbulos (COMT).
El ovocito secundario completa la meiosis II, forma un óvulo maduro y un segundo corpúsculo
polar. El núcleo del óvulo maduro se denomina pronúcleo femenino.
Los pronúcleos masculino y femenino se fusionan para crear un cigoto (una nueva célula cuyo
genotipo es una mezcla de cromosomas de origen materno y paterno).
Singamia es un término que describe el término exitoso de la fecundación, es decir, la formación
de un cigoto. La singamia se presenta cuando se fusionan los pronúcleos masculino y femenino y existe una maquinaria citoplasmática para una división celular apropiada.
La expectativa de vida de un cigoto es de sólo unas cuantas horas ya que su existencia termina con la primera división de la segmentación
SEGMENTACIÓN Y FORMACIÓN DEL BLASTOCISTO

La segmentación consiste en una serie de divisiones mitóticas del cigoto en la que el plano de la primera división pasa a través del área de la membrana plasmática en la que previamente habían sido expulsados los corpúsculos polares
La segmentación en humanos es holoblástica, lo que significa que las células se dividen por completo a través de su citoplasma. La segmentación en humanos es asimétrica, lo que significa que las células hijas no tienen un tamaño igual (es decir, una célula recibe más citoplasma que otra)
por lo menos durante las primeras divisiones celulares. La segmentación en humanos es asincrónica, lo que significa que sólo una célula se divide al mismo tiempo; por lo general, la célula hija más grande será la siguiente en dividirse por lo menos durante las primeras divisiones celulares.
El proceso de segmentación eventualmente forma una blástula que consiste en células llamadas blastómeros.
Un grupo de blastómeros (16 a 32 blastómeros) forma una mórula
Los blastómeros son totipotenciales hasta la etapa de ocho células (es decir, cada blastómero puede formar un embrión completo por si mismo). La totipotencialidad se refiere a que una célula madre se puede diferenciar en cualquier célula del organismo, incluidos los tejidos extraembrionarios.
Durante la formación del blastocisto se produce la secreción de líquido en el interior de la mórula que
ayuda a formar la cavidad del blastocisto. El producto se conoce ahora como blastocisto
La masa celular interna ahora se llama embrioblasto (que dará lugar al embrión). Las células del embrioblasto son pluripotenciales. La pluripotencialidad se refiere a que una célula madre se puede diferenciar en ectodermo, mesodermo y endodermo.
La masa celular externa ahora se llama trofoblasto (que dará lugar a la placenta).
La degeneración de la zona pelúcida se lleva a cabo el cuarto día después de la concepción. Esta zona debe degenerar para que se produzca la implantación.
IMPLANTACIÓN
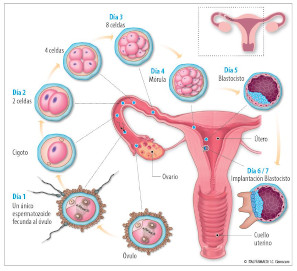
El blastocisto suele implantarse en la parte superior de la pared posterior del útero en el séptimo día después de la fecundación. La implantación tiene lugar en la capa funcional del endometrio durante la fase progestacional (secretora) del ciclo menstrual. El trofoblasto prolifera y se diferencia en citotrofoblasto y en sincitiotrofoblasto. El fracaso de la implantación puede implicar un rechazo inmunológico de la madre contra los antígenos del embrión (reacción del injerto contra el huésped)