
Гени
Ген – функціонально найменша одиниця генетичного апарату організму; транскрибована ділянка хромосоми, що кодує поліпептид, рРНК або тРНК. Дія генів проявляється у фенотипі. Термін запропонований В. Йогансеном в 1909 р. • Геном – сукупність генів гаплоїдного набору хромосом даного виду організмів.
Історія
У своєму розвитку генетика пройшла ряд етапів. Спадковістю люди цікавилися дуже давно. З розвитком сільського господарства сформувалася прикладна наука селекція, яка займалася створенням і формуванням нових порід тварин і сортів рослин. Проте, пояснити механізми передачі ознак нащадкам селекціонери не могли.
Уявлення про спадковість
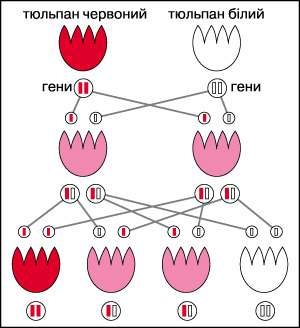
Спадковість — процес передачі в низці поколінь подібних ознак згідно з певними біологічними законами. Передача спадкових ознак і властивостей здійснюється в процесі розмноження. Основою розмноження є поділ клітин. В основі будь-якого виду поділу клітин, а значить і розмноження, лежить молекулярний механізм реплікації ДНК. Генетичний матеріал подвоюється, а потім рівномірно поділяється між дочірніми клітинами. При статевому розмноженні передача ознак здійснюється через гамети — яйцеклітини і сперматозоїди, при безстатевому — через соматичні клітини. Кожен вид організмів відтворює себе практично без змін з покоління в покоління протягом тисячоліть. В основі цього явища лежить висока стабільність спадкового матеріалу — молекул ДНК. Відомо багато закономірностей С. Цей процес залежить від багатьох причин, наприклад від того, в яких хромосомах знаходяться гени, скільки генів визначають ознаку, домінантні вони або рецесивні, зчеплені або незчеплені тощо.
Теорії
Ще до сучасної науки люди прагнули зрозуміти, як передаються риси від батьків до дітей. Спадковість – здатність організмів передавати наступному поколінню свої ознаки і властивості, тобто здатність відтворювати собі подібних.
Теорія часток
Гіпократ, відомий давньогрецький лікар, вважав, що кожна частина тіла батьків виробляє маленькі частинки, які впливають на розвиток потомства. Ці частинки (тепер відомі як "геммулки") потрапляли до статевих клітин і визначали риси дітей. Це була спроба пояснити спадковість через матеріальні компоненти, які передаються.
Спадкових рідин
Аристотель заперечував концепцію часток, вважаючи, що батьки передають своє потомству "спадкові рідини". Він стверджував, що ці рідини зливаються під час запліднення, створюючи новий організм. Це уявлення підкреслювало важливість обох батьків у формуванні потомства.
aПередформізм
У XVII-XVIII століттях виникла теорія передформізму, яка стверджувала, що в сперматозоїді або яйцеклітині вже міститься маленька, повністю сформована копія організму. Цей організм просто збільшувався у розмірах після запліднення. Це уявлення давало спрощене пояснення розвитку організмів, але не враховувало складність біологічних процесів. Ця концепція розвивалася протягом XVII-XVIII століть, зокрема, у роботах вчених, таких як Левенгук (1632–1723), який досліджував сперматозоїди та яйцеклітини.
Руйнування теорії
З розвитком біологічних наук і експериментів, зокрема роботи Карла Ернста фон Беара, було доведено, що розвиток організму відбувається поступово, починаючи з однієї клітини. Вчений встановив, що запліднена яйцеклітина не є "маленьким організмом", а має потенціал для розвитку, проходячи через стадії поділу і диференціації. Це спростувало теорію передформізму та відкрило нові шляхи для розуміння розвитку.
Пангенез
Чарльз Дарвін запропонував теорію пангінезису, згідно з якою маленькі частинки, які він назвав "геммулами", переносили спадкову інформацію від клітин тіла до статевих клітин. Дарвін вважав, що ці геммули були відповідальні за передачу риси потомству, і що їх кількість і якість могли змінюватися в залежності від впливу середовища.
Критика пангенезу
Ідеї Дарвіна піддавалися критиці, і вчені, такі як Грегор Мендель, згодом спростували концепцію пангінезису через свої експерименти на рослинах. Мендель встановив основи спадковості, формулюючи закони, які стали базою сучасної генетики. Це призвело до виникнення нових поглядів на спадковість, які враховували як хромосоми, так і гени, закладаючи основи для подальших досліджень у цій галузі.

Г. Мендель
Мендель Грегор Іоганн (20.07.1822, с. Хейнцендорф, Австрійської імперії – 1884) — видатний біолог, австрійський ботанік, засновник учення про спадковість.Захоплення біологією Мендель виявив дуже рано. Два роки він відвідував Інститут Олмюца, після цього став монахом в Августинському монастирі Святого Фоми. З 1844 до 1848 р. навчався в Богословському інституті у Брюнні. Але глибокі знання в багатьох галузях Мендель отримав завдяки самоосвіті. Нетривалий час Іоганн Грегор був викладачем, потім поїхав навчатися до Віденського університету. Саме там Мендель багато часу (1856–1863) присвятив дослідженням гібридних нащадків гороху, які стали складовою фундаменту генетики. Внаслідок своїх дослідів Мендель сформулював закони спадковості ознак у ряді поколінь (закони Менделя), створив наукові принципи опису та дослідження гібридів та їх нащадків (які саме форми брати до схрещування, як вести аналіз у першому та другому поколінні), розробив та застосував алгебраїчну систему символів та позначень ознак у генетиці.
a
Теорія спадковості
Хромосомна теорія спадковості — теорія, за якою матеріальними носіями спадковості є хромосоми. Це припущення було висловлено у 80–90 рр. XIX ст. після вивчення процесів поділу ядра клітини (мітозу і мейозу) та розкриття механізму збереження кількісної та якісної специфіки хромосомних наборів у наступних поколіннях.
Відкриття структури ДНК
Вотсон та Крік створили свою повну модель ДНК у перший тиждень березня 1953 року. Вся конструкція зійшлася як ідеально зібраний пазл. Коли її побачили Вілсон і Франклін, вони одразу підтвердили її правильність. 25 квітня 1953 року Вотсон та Крік опублікували статтю в журналі «Nature». В тому же номері вийшли статті Вілсона і Франклін, які підкріплювали модель експериментальними доказами. У 1962 році Вотсон, Крік та Вілкінс отримали за своє відкриття Нобелівську премію.
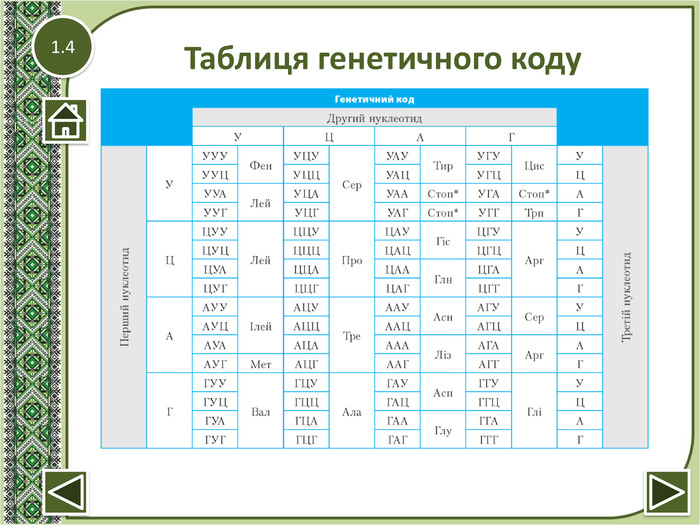
Генетичний код
Генетичний код — властива всім живим організмам єдина система запису спадкової інформації у вигляді послідовності нуклеотидів, в якій кожні три нуклеотиди (кодон) визначають одну молекулу амінокислоти у кодованому білку. Властивості генетичного коду: триплетність (3 нуклеотиди - 1 амінокислота), неперекривання (кодони одного гена не перекриваються), виродженість (кодування однієї амінокислоти кількома триплетами), компактність (код не містить нуклеотидів — проміжки між кодонами), універсальність (притаманний усім організмам).
aПроект "Геном людини"
Проект "Геном людини" - міжнародний проєкт наукових досліджень геному людини, мета якого полягає у визначенні послідовностей ДНК, локалізації генів та їх функцій.
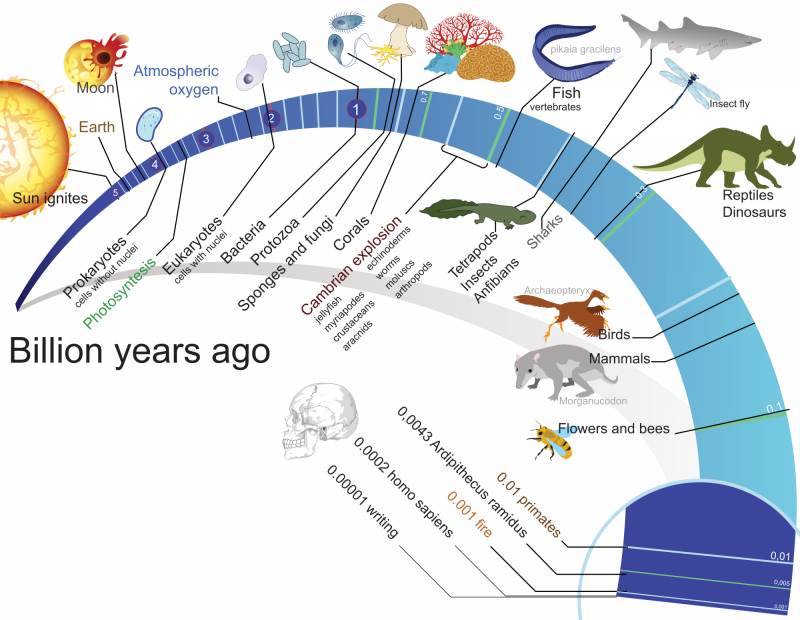
Еволюція
Еволюція — це процес зміни живих організмів у часі протягом історії Землі.

Еволюційна теорія
Теорія еволюції — наукова теорія, що пояснює механізми зміни форм живих організмів, їхніх спільнот та причини утворення біорізноманіття на Землі у процесі еволюції. Перші цілісні теорії еволюції були запропоновані на початку ХІХ сторіччя Еразмом Дарвіном та Жаном Батистом Ламарком.
Мутації
Мутації можна класифікувати за характером прояву, місцем виникнення, за характером змін спадкового апарату, за походженням тощо.

Природній добір
Природний добір — процес, в результаті якого виживають і залишають після себе потомство переважно особини з корисними в даних умовах змінами.Синтетична теорія еволюції розширила уявлення про цей процес. За СТЕ в результаті боротьби за існування виживають особини з найбільш корисними в даному середовищі фенотипами. Якщо відбір фенотипів триває у ряду поколінь, то в популяції відбудеться зміна генофонду.Природний добір — спрямований процес вибіркового збереження фенотипів і відтворення адаптивних генотипів.Природний добір — це механізм, за допомогою якого живі організми адаптуються і еволюціонують. Окремі організми, найкраще пристосовані до середовища, виживають і успішно розмножуються, народжуючи плідне потомство. Після численних циклів розмноження такі організми стають переважаючими в популяції. У результаті формується пристосованість до умов середовища.
a
Роль генів в еволюції
Гени відіграють ключову роль у процесі еволюції, оскільки вони є носіями спадкової інформації, яка передається від одного покоління до наступного. В еволюційному контексті гени визначають, які риси і ознаки будуть передані потомству, а також яким чином популяції організмів будуть змінюватися під впливом природного відбору та інших еволюційних механізмів.
a
Адаптація
Генотипічна адаптація — відбір спадково детермінованої (зміна генотипу) підвищеної пристосованості до змінених умов (спонтанний мутагенез) фенотипічна адаптація — при цьому відборі мінливість обмежена нормою реакції, визначуваної стабільним генотипом.
a
Поліморфізм
Поліморфізм - це генетичний варіант, який з'являється щонайменше у 1% населення. (наприклад, групи крові АВО людини, резус-фактор людини та основний комплекс гістосумісності людини). Встановлюючи відсічення на рівні 1%, він виключає спонтанні мутації, які могли статися в - і поширюватися через нащадків - однієї сім'ї.
Генетичні зміни
Генетичні зміни — це процеси, які призводять до змін у послідовності ДНК організму. Вони відіграють ключову роль в еволюції та адаптації організмів, а також є основою для виникнення нових рис і захворювань. Основними джерелами генетичних змін є мутації та генетична рекомбінація. Обидва механізми забезпечують генетичну різноманітність у популяціях і є важливими факторами еволюційних процесів.

Через мутацію
Мутації — це зміни в нуклеотидній послідовності ДНК. Вони можуть виникати через помилки в реплікації ДНК, вплив радіації, хімічних речовин або вірусів. Мутації можуть бути спадковими (якщо вони відбулися в статевих клітинах) або соматичними (якщо вони виникають у клітинах тіла, що не передаються потомству).Мутації можуть виникати:Спонтанно — під час нормальних процесів клітинного поділу, коли ДНК не завжди реплікується точно.Індуковані мутації — викликані зовнішніми факторами, такими як ультрафіолетове випромінювання, радіація або хімічні речовини.Наслідки мутацій.Позитивні мутації можуть давати організму еволюційні переваги, сприяючи адаптації до середовища.Негативні мутації можуть бути шкідливими, викликаючи захворювання або дисфункції.Нейтральні мутації не впливають на функціонування організму і залишаються в геномі без явних наслідків.Мутації створюють нові варіанти генів, а рекомбінація забезпечує їх розповсюдження у популяції, створюючи різноманітні комбінації цих варіантів. Взаємодія цих процесів забезпечує основу для еволюційних змін і адаптації організмів до нових умов середовища.

Через рекомбінацію
Генетична рекомбінація — це процес обміну генетичним матеріалом між двома хромосомами, що забезпечує нові комбінації генів у потомства. Вона відбувається під час мейозу (утворення статевих клітин) і є основним джерелом генетичної різноманітності у організмів із статевим розмноження. КросинговерЦе процес обміну сегментами між парними (гомологічними) хромосомами під час мейозу. Внаслідок кросинговеру утворюються нові комбінації алелів, що відрізняються від батьківських. Це дозволяє потомству мати унікальні комбінації генів.Незалежне комбінування хромосомКоли гамети утворюються під час мейозу, хромосоми від матері і батька розходяться незалежно одна від одної. Це означає, що кожна гамета містить випадковий набір материнських і батьківських хромосом. Це також забезпечує генетичну різноманітність.Алельна рекомбінаціяРекомбінація може відбуватися не лише між цілими хромосомами, а й між окремими генами або їх частинами. Це призводить до появи нових алельних комбінацій, які можуть змінити функцію або регуляцію генів.Генетичні конверсіїГенетична конверсія — це специфічний вид рекомбінації, коли одна алель змінюється на іншу через обмін сегментами між гомологічними хромосомами. Це явище допомагає створювати нові генетичні варіанти в популяціях.Наслідки рекомбінаціїПідвищення генетичної різноманітності. Завдяки рекомбінації, потомство отримує унікальні комбінації генів, що сприяє адаптації і виживанню в умовах змін навколишнього середовища.Захист від шкідливих мутацій. Якщо одна з батьківських хромосом містить мутацію, рекомбінація може створити гамети, які будуть містити лише здорові варіанти генів.Еволюційні переваги. Нові комбінації генів можуть призводити до появи нових ознак або покращувати наявні властивості, що надає організмам еволюційні переваги.

Транспозони
Транспозони - це сегменти ДНК, які можуть переміщатися в різні положення в геномі однієї клітини. У процесі вони можуть викликати мутації і збільшувати (або зменшувати) кількість ДНК в геномі клітини, а якщо клітина є попередником гамети, в геномах будь-яких нащадків. Ці рухливі сегменти ДНК іноді називають «стрибаючими генами», і існує два різних типи. Транспозони II класу складаються з ДНК, яка рухається безпосередньо з місця на місце. Транспозони класу I - це ретротранспозони, які спочатку транскрибують ДНК в РНК, а потім використовують зворотну транскриптазу, щоб зробити копію ДНК РНК для вставки в нове місце.

Транспозони I класу
Ретротранспозони також рухаються механізмом «копіювання та вставки», але на відміну від описаних вище транспозонів, копія зроблена з РНК, а не ДНК. Копії РНК потім транскрибуються назад в ДНК - за допомогою зворотної транскриптази - і вони вставляються в нові місця в геномі. Багато ретротранспозонів мають довгі термінальні повтори (LTR) на своїх кінцях, які можуть містити понад 1000 базових пар у кожному. Як і транспозони ДНК, ретротранспозони генерують прямі повтори на своїх нових місцях вставки. Насправді саме наявність цих прямих повторів часто є підказкою про те, що втручається ділянку ДНК прибув туди шляхом ретротранспозиції. Близько 50% всього генома людини складається з ретротранспозонів.
Транспозони ІІ класу
Транспозони класу II переміщуються за допомогою процесу «вирізати і вставити»: транспозон вирізається з його розташування (як Command/Control-X на вашому комп'ютері) і вставляється в нове місце (Command/Control-V). Цей процес вимагає ферменту - транспозази - який кодується в межах деяких з цих транспозонів.
Вплив
Транспозони мають значний вплив на генетичну варіативність і еволюцію, оскільки вони здатні переміщатися в межах геному, змінюючи його структуру та функцію.
Варіативність
Вставки транспозонів у гени можуть призвести до мутацій, що змінюють функцію гена. Наприклад, якщо транспозон вставляється у регуляторну або кодувальну область гена, він може порушити або повністю зупинити його функціонування.Це призводить до появи нових алелів, які можуть бути нейтральними, шкідливими або корисними для організму, залежно від контексту.Переміщення та делеціїТранспозони можуть переміщувати великі фрагменти геному, що створює структурні варіації, такі як дуплікації, інверсії, інсерції та делеції. Такі зміни можуть впливати на кількість і структуру генів.Наприклад, дуплікація гена, викликана транспозонами, може створювати умови для функціональної дивергенції генів, що з часом веде до нових біологічних функцій.Генетичні рекомбінаціїТранспозони можуть виступати як сайти для рекомбінації, що дозволяє їм сприяти обміну генетичним матеріалом між різними частинами геному або навіть між різними організмами. Така рекомбінація підвищує рівень генетичної різноманітності та адаптації.
Еволюція
Геномна еволюціяТранспозони є основними двигунами геномної еволюції. Їхня здатність переміщуватися та спричиняти геномні зміни відіграє важливу роль у формуванні нових структурних варіацій геному, таких як збільшення його розміру або створення нових функціональних елементів.У рослин і тварин транспозони можуть становити значну частину геному (до 90% у деяких видів). Наприклад, геном кукурудзи складається майже на 85% з транспозонів, що підкреслює їхній значний вплив на еволюцію цього виду.Створення нових генівРетроелементи (ретротранспозони) можуть копіювати не лише себе, але й прилеглі послідовності ДНК, включаючи гени, що створює нові копії генів в інших частинах геному. Це відкриває можливості для появи нових функцій у організмів.Такі нові гени можуть бути підхоплені природним добором, що забезпечує еволюційні переваги для виду.Еволюційна адаптаціяТранспозони можуть вносити адаптивні зміни, які можуть допомогти організмам вижити в змінюваних умовах. Наприклад, в деяких випадках транспозони можуть регулювати експресію генів, які відповідають за стійкість до екологічних стресорів, таких як засуха або патогени.Вони можуть також впливати на розвиток адаптивного імунітету, наприклад, через модифікацію генів, що відповідають за захист від вірусів.Ко-еволюція з геномомГеноми організмів і транспозони знаходяться в постійному процесі ко-еволюції. Організми розвивають механізми, які регулюють і обмежують активність транспозонів, оскільки неконтрольоване переміщення транспозонів може бути шкідливим. Наприклад, у багатьох видів існують механізми епігенетичного контролю, які вимикають транспозони через метилювання ДНК або модифікацію хроматину.Проте транспозони постійно знаходять нові способи уникнути цих механізмів регуляції, що створює динамічний баланс між геномом і транспозонами.

Структура
Гени класифікують:за розташуванням у клітинах: ядерні та цитоплазматичні гени (розташовані в мітохондріях і хлоропластах);за функціональним значенням: структурні (кодують білок або РНК) та регуляторні (визначають початок, швидкість та послідовність процесів синтезу РНК на матриці ДНК.за функціональним продуктом експресії: білкові гени, які містять інформацію про первинну структуру білка (на основі цих генів для біосинтезу утворюються іРНК або матричні мРНК); та РНК-гени, які кодують велику кількість різних видів РНК та не піддаються трансляції (це тРНК, рРНК, маленькі ядерні мяРНК, мікро-РНК);за активністю: конститутивні гени — це гени, що є постійно активними, осільки білки, які ними кодуються необхідні для постійної клітинної діяльності. Ці гени забезпечують синтез білків, ферментів і не потребують спеціальної регуляції. Неконститутивні гени — це гени, які стають активними лише тоді, коли білок, який вони кодують потрібен клітини. Вони регулюються клітиною або організмом і за участі кодованих білків забезпечують диференціацію клітин.
Гени
Ген складається з ДНК, яка «читається» або транскрибується для отримання молекули РНК під час процесу транскрипції. Один з основних типів молекули РНК, званий месенджерної РНК (мРНК), надає інформацію для рибосоми для каталізації синтезу білка в процесі, який називається трансляцією. Процеси транскрипції і перекладу в сукупності називають експресією генів. Експресія генів - це синтез специфічного білка з послідовністю амінокислот, яка закодована в гені. Потік генетичної інформації від ДНК до РНК до білка описується центральною догмою. Ця центральна догма молекулярної біології додатково з'ясовує механізм гіпотези Бідл та Татума «один ген-один фермент». Кожен з процесів реплікації, транскрипції і трансляції включає етапи 1) ініціації, 2) подовження (полімеризації) і 3) припинення.
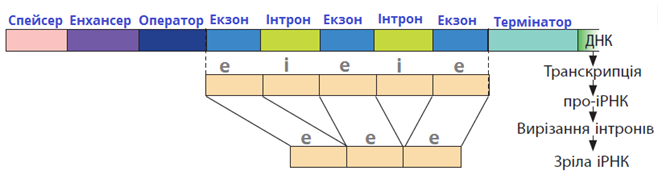
Екзони
Багато екзонів кодують одиницю, дуже близьку до білкового домену, наприклад екзони леггемоглобіну, або змінні та постійні області імуноглобулінів або доменів (наприклад, «kringle») у попереднику EGF, які також знаходяться в частині рецептора ЛПНЩ. Організація екзонів, як правило, добре зберігається у сильно розбіжних видах. Інтрони, як правило, виникають між тими частинами генів, які кодують структурні домени білків. Дублювання екзонів, що кодують структурні домени і подальша рекомбінація може призвести до більш швидкої еволюції нового білка, по суті використовуючи частини з раніше еволюціонували генів. Аналогічно будівництву будинку зі збірних деталей, на відміну від одного цвяха і однієї дошки одночасно — почніть з попередньо зібраних стін, балок даху тощо.Однак взаємозв'язок між екзонами та структурними доменами білків не є точним, і деякі межі екзон-інтронів змінюються (трохи) у генах для різних видів. Інша модель стверджує, що інтрони є транспозіруемимі елементами. Вони можуть вставляти будь-де в ген, але вони найменш руйнівні в межах домену, і ці останні вставки, швидше за все, фіксуються в популяції, ніж вставки в середину області, що кодує домен. Отже, результати після довгих років еволюції полягають у тому, що інтрони, як правило, знаходяться між доменами кодування області, але ген спочатку був неушкодженим, а не зібраним з дискретних екзонів.
Інтрони
Інтро́н — ділянка ДНК, яка є частиною гену, але на відміну від екзонів, не містить інформації про послідовність амінокислот білка. Хоча інтрони й не несуть інформації про послідовність амінокислот у білку, вони іноді дозволяють альтернативний сплайсинг гену, таким чином, що розділені інтроном ділянки можуть комбінуватися у різній послідовності, виробляючи різні варіанти білка з одного гену. Контроль сплайсингу мРНК і, в результаті, вибір кінцевого продукту, виконується широкою різноманітністю сигнальних молекул. Інтрони також іноді містять «застарілий код» секції гена, що ймовірно колись був частиною білка, але потім перестав використовуватися.Загалом вважається, що послідовність інтронів є ділянкою ДНК без певної функції. Проте зараз це викликає сумнів: відомо, що інтрони містять декілька коротких послідовностей, які важливі для ефективного сплайсингу. Точний механізм дії цих інтронних енхансерів ще недостатньо відомий, але вважається, що вони служать обов'язковими ділянками ефективного синтезу білків, що стабілізують сплайсосому. Також можливо, що вторинна структура РНК, сформована з використанням послідовності інтронів, також має ефект на сплайсинг, інтронна послідовність при одному альтернативному сплайсингу стає екзонною при іншому. Послідовності «застарілого коду», з другого боку, в більшості випадків дійсно є «еволюційними артефактами».
a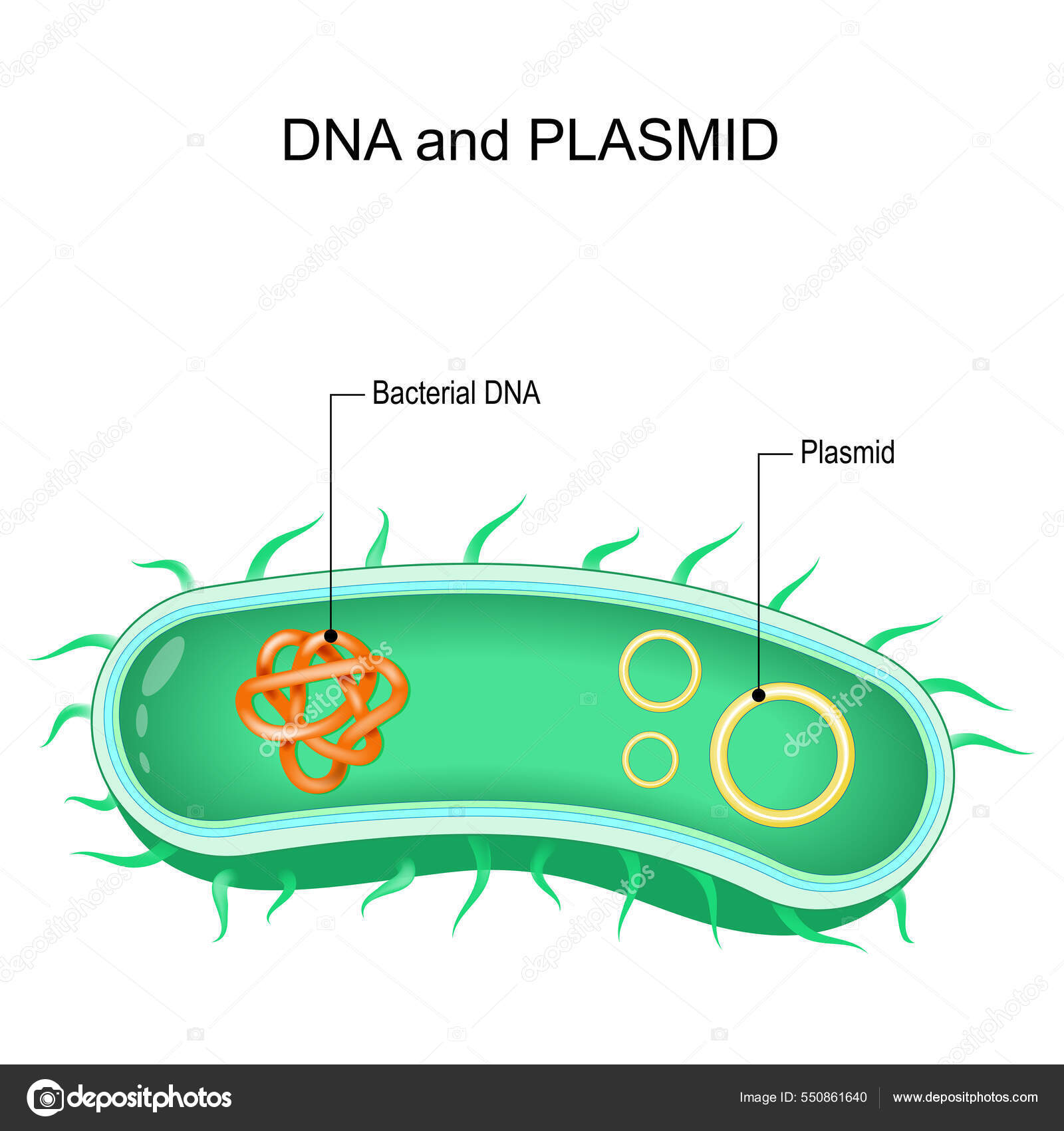
Плазміди
Плазміди - фрагменти ДНК, позахромосомні (додаткові по відношенню до хромосоми) генетичні структури бактерій, здатні автономно розмножуватися і існувати в цитоплазмі бактеріальної клітини. Виділяють автономні (не пов'язані з хромосомою бактерії) та інтегровані (вбудовані в хромосому) плазміди. Термін «плазміда» введений американським генетиком Ледербергом в 1952 р. для позначення статевого фактора бактерій (F-фактора, F-плазміди), виявленого в клітинах культури кишкової палички та відповідального за їх здатність бути донорами генетичного матеріалу (молекул ДНК) при кон'югації з клітинами-реципієнтами, не містять статевого фактора.Автономні плазміди існують в цитоплазмі бактерій і здатні самостійно репродукуватися; в клітині може бути присутнім кілька їхніх копій.Інтегровані плазміди репродукуються одночасно з бактеріальною хромосомою. Інтеграція плазмід відбувається при наявності гомологічних послідовностей ДНК, при яких можлива рекомбінація хромосомної та плазмідної ДНК .Плазміди також поділяють на трансмісивні (наприклад, F-або R-плазміди), здатні передаватися за допомогою кон'югації, і не трансмісивні.Плазміди виконують регуляторні або кодуючи функції. Регуляторні плазміди беруть участь в компенсуванні тих чи інших дефектів метаболізму бактеріальної клітини за допомогою вбудовування в пошкоджений геном і відновлення його функцій. Плазміди приносять в бактеріальну клітину нову генетичну інформацію, що кодує нові, незвичайні властивості (наприклад, стійкість до антибіотиків).
Роль
Плазміди часто містять гени, які не є необхідними для базових життєвих функцій бактерій, але дають їм переваги в певних умовах:Стійкість до антибіотиків. Один з найпоширеніших прикладів — плазміди, що містять гени стійкості до антибіотиків. Це дозволяє бактеріям виживати в присутності антибіотиків, які інакше їх би знищили. Такі плазміди передаються між бактеріями і можуть швидко поширювати стійкість у популяції.Стійкість до важких металів. Деякі плазміди містять гени, які дозволяють бактеріям виживати в середовищах, забруднених токсичними металами, такими як ртуть, кадмій або мідь.Фактори вірулентності. Плазміди можуть нести гени, які надають бактеріям здатність викликати хвороби у рослин, тварин або людей. Такі плазміди відіграють важливу роль у патогенезі.Плазміди також сприяють різноманітності генетичного матеріалу в популяціях бактерій, оскільки вони можуть переносити гени з різних джерел.
Плазмідна ДНК
Плазмідна ДНК — це невеликі кільцеві молекули ДНК, що знаходяться поза основною хромосомною ДНК у клітинах, переважно у прокаріотів (бактерій), але іноді зустрічаються і у деяких еукаріотів (наприклад, у дріжджів). Плазміди можуть реплікуватися автономно і виконують низку важливих функцій для клітин, включаючи:Передача корисних генів. Вони часто містять гени, що надають бактеріям певні переваги, наприклад, стійкість до антибіотиків, токсичних речовин або захист від фагів.Генетична різноманітність. Плазміди сприяють генетичній варіативності шляхом передачі генів від однієї клітини до іншої, навіть між різними видами бактерій.Горизонтальний генний перенос. Плазміди є ключовими учасниками кон’югації, одного з механізмів горизонтального переносу генетичного матеріалу між клітинами.
Рекомбінантні плазміди
Рекомбінантні плазміди створюються у лабораторії шляхом злиття генетичного матеріалу з різних джерел. Вони використовуються як інструменти для експериментів у галузі генетики та молекулярної біології. Ось як працює створення рекомбінантних плазмід:Створення рекомбінантних плазмідВставка чужорідного гена. Для створення рекомбінантної плазміди використовують ферменти рестриктази, які розрізають плазмідну ДНК у визначених місцях, і лігази, що з’єднують чужорідну ДНК з плазмідною ДНК. У цей процес може бути вставлений будь-який генетичний матеріал, наприклад, гени, що кодують білки.Клонування генів. Після того як чужорідний ген вставляється в плазміду, цю конструкцію можна вводити у бактерії (зазвичай у Escherichia coli), де плазміда буде реплікуватися разом з бактеріальною ДНК, забезпечуючи багатократне копіювання вставленого гена.Застосування рекомбінантних плазмідВиробництво рекомбінантних білків. За допомогою рекомбінантних плазмід бактерії можуть виробляти білки, що кодуються чужорідними генами. Це важливо для отримання таких білків, як інсулін, інтерферони або фактори згортання крові.Генна терапія. Рекомбінантні плазміди використовуються для введення нормальних копій генів у клітини пацієнтів з генетичними захворюваннями.Вакцини на основі ДНК. Рекомбінантні плазміди можуть містити гени, що кодують антигени патогенів, які, після введення в організм, стимулюють імунну відповідь і захищають від інфекцій.Вивчення функцій генів. Дослідники використовують рекомбінантні плазміди для з’ясування ролі різних генів та їх взаємодій у клітинах.ПеревагиЛегкість маніпуляції. Плазміди мають невеликий розмір, тому з ними легко працювати під час молекулярних експериментів.Автономна реплікація. Рекомбінантні плазміди можуть реплікуватися незалежно від хромосомної ДНК, що дозволяє легко отримати велику кількістьвводячи в них певні гени, що дозволяє точно вивчати їх функції або використовувати їх для терапевтичних цілей.ПрикладиpBR322 — один із найперших векторів для клонування, використовуваний для введення чужорідних генів у бактерії.pUC плазміди — широко використовувані вектори в біотехнології для швидкого й ефективного клонування генів.
Мобільні генетичні елементи
Мобільні елементи геному — послідовності ДНК, здатні переміщатися усередині геному живих організмів. Існує декілька класів мобільних елементів геному, що відрізняються за будовою і способом переміщення:Інсерційні елементи, наприклад, IS1603Транспозони, наприклад, Tn5 — латентна форма помірних бактеріофагів, наприклад, транспозонподібного фагу Mu (мю)Плазміди, наприклад, статеві ворсинки кишкової палички F (еф)Хоча мобільні елементи в цілому є «генетичними паразитами», викликаючи мутації в генетичному матеріалі організму хазяїна, і знижуючи його пристосованість за рахунок витрати ресурсів на реплікацію і синтез білків паразиту, вони є важливим механізмом мінливості і обміну генетичним матеріалом між організмами як одного виду, так і різних видів.

Транспозони
Нове дослідження під керівництвом Університету Південної Каліфорнії підкреслює, що транспозони, які здатні переміщатися в різні частини геному, пов’язані з віковими захворюваннями. Ці ділянки, які ще називають «генами, що стрибають», становлять приблизно 45 % ДНК людини, і їхня активність значною мірою пригнічена в молодших і більш здорових клітинах. Проте з віком ці гени проявляються більше і стають мобільнішими, що корелює з різними віковими дисфункціями. Головне питання вчених полягало в тому, чи є підвищена активність транспозонів лише побічним продуктом старіння, або вони, навпаки, є причиною вікових змін.
Відкриття
Історія відкриття транспозонів є важливою віхою у розумінні генетичних елементів та їхньої ролі у геномі. Транспозони (або мобільні генетичні елементи) — це сегменти ДНК, які можуть змінювати своє положення в межах геному, переміщуючись з однієї частини ДНК до іншої. Відкриття транспозонів пов’язане з дослідженнями американської вченої Барбари МакКлінток, яка працювала на моделі кукурудзи. У 1940-х роках вона вивчала спадкові зміни в кольорі зерен кукурудзи та виявила незвичайні явища:МакКлінток помітила, що деякі гени, відповідальні за колір зерен кукурудзи, можуть раптово змінювати своє положення у геномі.Вона також звернула увагу на те, що ці зміни в кольорі не були постійними, а могли "перемикатися" у наступних поколіннях.На основі цих спостережень МакКлінток висунула ідею про те, що в геномі існують мобільні елементи, які можуть пересуватися між різними ділянками ДНК, викликаючи генетичні зміни:Вона назвала ці елементи "контролюючими елементами".МакКлінток вважала, що переміщення таких елементів може призводити до мутацій і змін у експресії генів.Спочатку роботи МакКлінток не були прийняті науковою спільнотою з ентузіазмом. Її ідеї про мобільні елементи здавалися революційними і суперечили традиційним уявленням про стабільність генетичного матеріалу.У той час науковці вважали, що гени мають постійне місце у хромосомах і не можуть переміщатися.У 1960-х і 1970-х роках, коли молекулярна біологія активно розвивалася, вчені підтвердили ідеї МакКлінток про мобільні генетичні елементи. Було відкрито багато транспозонів у різних організмів, таких як бактерії, дріжджі, рослини і тварини:Дослідження на бактеріях, зокрема на Escherichia coli, підтвердили існування мобільних елементів, які можуть переносити гени стійкості до антибіотиків.У результаті цих досліджень стало зрозуміло, що транспозони є важливими елементами еволюції і можуть відігравати ключову роль у генетичній варіативності.За своє революційне відкриття транспозонів Барбара МакКлінток отримала Нобелівську премію з фізіології або медицини у 1983 році.Її відкриття допомогло зрозуміти механізми мутацій і вплив транспозонів на еволюцію.Це також привело до нових досліджень у галузі генетики, зокрема вивчення регуляції генів і геномної пластичності.
Функції
Транспозони мають кілька ключових функцій у геномі, які впливають на його структуру та функцію:Геномна пластичність. Транспозони забезпечують мобільність генетичного матеріалу, змінюючи структуру геному через переміщення або копіювання себе.Генетична рекомбінація. Переміщення транспозонів може сприяти генетичній рекомбінації, що може змінювати експресію генів або створювати нові алелі.Мутації. Транспозони можуть викликати мутації через вставки у функціональні гени або регуляторні ділянки, що може призводити до втрати або зміни функцій генів.Регуляція експресії генів. Деякі транспозони здатні діяти як елементи регуляції генів, активуючи або блокуючи їхню експресію.Еволюція геному. Через постійні переміщення транспозони сприяють еволюційним змінам у геномах еукаріотів, додаючи генетичне різноманіття.

Представники
Як відомо із попередньо переглянутої інформації, транспозони є:В еукаріотів транспозони поділяються на два основні класи:Транспозони І класу (ретротранспозони):Лінійні (LINEs): автономні транспозони, які здатні копіювати та вставляти себе за допомогою механізму зворотної транскрипції.Короткі повтори (SINEs): неавтономні транспозони, що використовують білки, синтезовані LINE-ами для транспозиції.LTR-ретротранспозони: містять довгі повтори та за своєю структурою подібні до ретровірусів.Транспозони ІІ класу:Транспозони типу "виріж і встав": переміщуються за допомогою ферменту транспозази.Ac/Ds (Activator/Dissociation): мобільні елементи, виявлені у кукурудзи Барбарою МакКлінток.

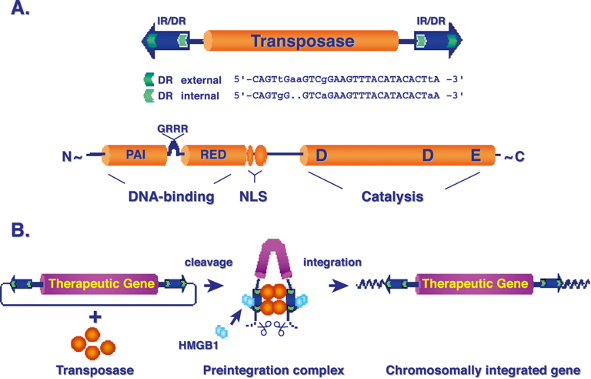
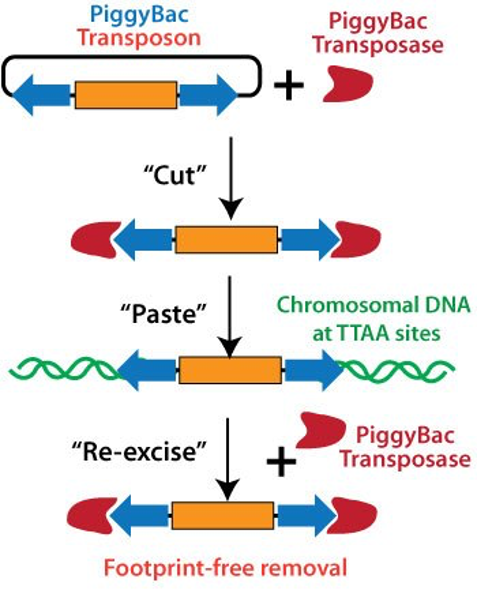
ДНК-транспозони
ДНК-транспозони переміщуються за допомогою ферменту транспозази, який вирізає транспозон з його початкового місця та вставляє в нове місце в геномі.Вони зазвичай мають інвертовані повтори (IR, inverted repeats) на кінцях, які є важливими для процесу переміщення. Кодуючий регіон транспозона часто містить ген транспозази – ферменту, який і відповідає за їх рух.

Tn5 і Tn10
Транспозон Tn5 – це бактеріальний ДНК-транспозон, який має велике значення для досліджень в галузі генетики та молекулярної біології. Tn5 використовується для створення мутацій і переміщення генетичних елементів у бактерій.Структура Tn5:Tn5 містить ген транспозази, яка відповідає за його мобільність.Має інвертовані повтори на кінцях, що необхідні для процесу транспозиції.У складі Tn5 є кілька генів стійкості до антибіотиків (наприклад, стійкість до канаміцину та неоміцину), що часто використовують для відбору мутантів у лабораторних умовах.Механізм переміщення:Tn5 рухається за механізмом "вирізати-вставити" за допомогою транспозази.Транспозаза розпізнає інвертовані повтори на кінцях транспозона, вирізає транспозон і вставляє його в нове місце в геномі.Застосування:Використовується як інструмент для мутагенезу в бактеріях.Допомагає вивчати генетичні регуляторні елементи через вставки транспозона у різні ділянки геному.Tn10Транспозон Tn10 – також бактеріальний ДНК-транспозон, схожий на Tn5, але з власними особливостями.Структура Tn10:Tn10 містить ген транспозази, необхідний для його мобільності, і також має інвертовані повтори на кінцях.Включає ген, що надає стійкість до тетрацикліну – це один із найпоширеніших антибіотиків, що використовується для створення селекційного тиску в бактеріях.Механізм переміщення:Транспозиція Tn10 відбувається також за принципом "вирізати-вставити", але на неї може впливати стан клітини, наприклад, рівень ДНК-реплікації.Транспозон може індукувати мутації, вставляючись у геном і порушуючи функції генів.Застосування:Tn10 використовується для вивчення бактеріальної генетики, включаючи дослідження регуляції генів і механізмів антибіотикорезистентності.Його інсерції можуть порушувати гени та дозволяють досліджувати функціональну генетику бактерій.Важливість Tn5 і Tn10:Ці транспозони є важливими інструментами для генетичних досліджень, дозволяючи вивчати структуру генів, геномну організацію, і навіть використовувати їх у практичній біотехнології для створення генетичних модифікацій.
aAc/Ds елементи
Елемент Ac був відкритий Барбарою МакКлінток під час вивчення кукурудзи. Це автономний ДНК-транспозон, тобто він містить усі гени, необхідні для транспозиції, включаючи транспозазу.Структура Ac:Містить ген транспозази, який відповідає за переміщення як самого Ac, так і пов'язаних елементів Ds (дисоціаторів).Має інвертовані повтори на своїх кінцях, що є ключовими для транспозиції.Функція Ac:Ac здатен самостійно переміщуватися у геномі, а також може ініціювати переміщення неавтономних транспозонів (як-от Ds).Елемент Ds є неавтономним транспозоном, що означає, що він не містить свого власного гена транспозази та потребує Ac для переміщення.Структура Ds:Не має активного гена транспозази, тому самостійно не може транспонуватися.Має інвертовані повтори на кінцях, схожі до Ac, які розпізнає транспозаза Ac.Функція Ds:Коли Ac присутній у геномі, Ds може транспонуватися, що може призвести до мутацій або порушень у експресії генів.Часто вставка Ds у ген може викликати зміни фенотипу, наприклад, зміну кольору зерен кукурудзи.Значення Ac/Ds у генетиці:Відкриття Ac/Ds елементів допомогло зрозуміти, що геном є динамічним і може змінюватися через мобільні генетичні елементи.Ac/Ds елементи використовувалися для вивчення рекомбінації та генетичної регуляції у рослин і зробили великий внесок у розвиток генетики мобільних елементів.
a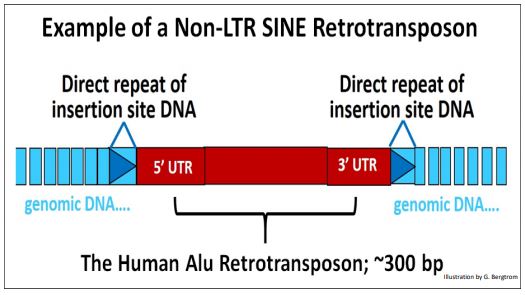
Ретротранспозони
Ретротранспозони — це мобільні генетичні елементи, що застосовують метод «копіювати й вставити» для розповсюдження в геномі тварин.
a
LTR-ретротранспозони
LTR-ретротранспозони є мобільними генетичними елементами, що копіюють себе через механізм, подібний до ретровірусів. Їхнім характерним елементом є довгі кінцеві повтори (LTR), які знаходяться на обох кінцях транспозона.Структура:LTR: Повторювані послідовності на кінцях транспозона, які містять сигнали для транскрипції та інтеграції в геном.Гени: Містять гени, що кодують зворотну транскриптазу (для створення ДНК з РНК) і інтегразу (для інтеграції в геном).Механізм:Транспозон спочатку транскрибується в РНК, після чого зворотна транскриптаза перетворює цю РНК назад у ДНК.Інтеграза вставляє нову копію транспозона в інше місце геному.

Non-LTR-ретротранспозони
Non-LTR-ретротранспозони не містять довгих кінцевих повторів (LTR), але також копіюються через механізм зворотної транскрипції. Ці транспозони є одними з найпоширеніших у геномах багатьох організмів.Структура:Відсутність LTR-послідовностей.Включають гени, що кодують зворотну транскриптазу та ендонуклеазу для копіювання та вставки.Механізм:Non-LTR-ретротранспозони використовують зворотну транскриптазу для копіювання РНК у ДНК, але механізм інтеграції у геном менш організований порівняно з LTR-ретротранспозонами.
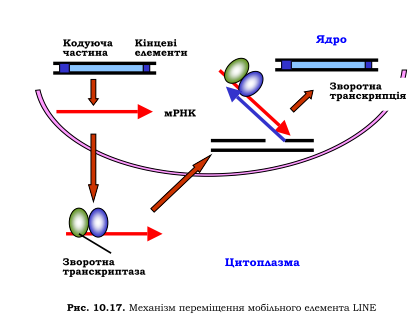
LINEs
LINEs — довгі (кілька тисяч пар заснованих) послідовності ДНК у геномі еукаріотів, що представляють собою ретротранспозони, не містять довгих концевих повторів.Перший опис LINE-послідовності, довжиною близько 6 400 пар, було опубліковано в 1980 році.
LINE-1 (L1)
LINE1 (абревіатура від Long interspersed ядерний елемент-1, також відомий як L1 і LINE-1) — це сімейство споріднених переміщуваних елементів класу I в ДНК багатьох груп еукаріот, включаючи тварин і рослини, класифікованих як довге вкраплене ядро елементи (ЛІНІЇ). Транспозони L1 найбільш поширені серед ссавців, де вони становлять значну частку загальної довжини геному, наприклад, вони складають приблизно 17% геному людини.
a
SINEs
SINEs - короткі послідовності ДНК (менше 500 пар заснованих) в геномі еукаріот, що з'явилися в результаті зворотної транскрипції коротких молекул РНК, транскрибованих РНК-полімеразою III: 5S рРНК, тРНК і різні мяРНК.SINE не кодують білки, і їх транспозиція в геном залежить від інших мобільних елементів.Розрізняють:SINE, що виникли від тРНК (звичайні у беспозвоночных і позвоночных тварин, у багатьох квіткових рослин);SINE, що виникли від 7SL РНК (встречаются тільки у гризунів, приматів і тупаїв);SINE, що виникли від 5S рРНК (знайдені у деяких риб і у деяких молокопитающих).
Alu-повтори
Alu-повтор, Alu-елемент - коротка послідовність ДНК, яка була відкрита при обробці ДНК людини рестриктазою Alu. Alu-повтор належить до класу коротких диспергованих повторів (SINEs). Alu-повтори різних типів у великій кількості представлені у геномах приматів. У людському геномі є одними з найпоширеніших елементів. Alu-повтор походить від гена, що кодує 7SL РНК, яка є компонентом сигналу розпізнаючої частинки.
МІТЕ
МІТЕ – це короткі неавтономні ДНК-транспозони, які не містять генів, необхідних для їх мобільності. Вони зазвичай мають інвертовані повтори на кінцях і потребують допомоги інших автономних транспозонів для переміщення.Структура:Малі транспозони з інвертованими кінцевими повторами.Відсутні гени, що кодують транспозазу.Механізм:МІТЕ потребують допомоги від автономних транспозонів, які надають необхідну транспозазу для переміщення.
a
Tourist
Tourist — це тип МІТЕ (Miniature Inverted-repeat Transposable Elements), невеликих неавтономних транспозонів, поширених у рослинах. Вони є короткими ДНК-послідовностями з інвертованими кінцевими повторами, які потребують допомоги автономних транспозонів для переміщення.Характеристики:Вони не містять генів для транспозиції, таких як транспозаза, тому не можуть самостійно переміщатися.Tourist має інвертовані кінцеві повтори і може сприяти геномній мінливості.Механізм:Ці елементи можуть переміщуватися в геномі за допомогою транспозази, що кодується автономними транспозонами. МІТЕ, такі як Tourist, часто зустрічаються в кодуючих ділянках генів і можуть впливати на експресію генів.Приклади:Tourist елементи зустрічаються у різних рослинних геномах, таких як кукурудза, рис, пшениця.
Stowaway
Stowaway — це ще один приклад МІТЕ елементів, що подібні до Tourist. Вони також є неавтономними і можуть переміщуватися в геномі за допомогою транспозази від інших транспозонів.Характеристики:Як і Tourist, вони мають короткі інвертовані кінцеві повтори, але не кодують жодного білка для самостійного переміщення.Stowaway елементи можуть впливати на структуру та функції генів.Механізм:Переміщення Stowaway елементів залежить від присутності транспозази автономних транспозонів. Вони здатні вставлятися в різні ділянки геному, впливаючи на генетичну мінливість і регуляцію генів.Приклади:Знайдені у багатьох рослинах, наприклад, у рисі та інших злакових культурах.
Консервативні
Консервативні транспозони переміщуються з однієї ділянки геному в іншу без збільшення кількості копій. Це відбувається за механізмом "вирізати-вставити", коли транспозон вирізається з одного місця і вставляється в інше.Структура:Містять гени, що кодують транспозазу.Мають інвертовані повтори на кінцях.Механізм:Переміщення за принципом "вирізати-вставити", тобто транспозон вирізається з одного місця і вставляється в інше.
a
Tn3 у бактерій
Tn3 — це приклад транспозона у бактерій, який належить до класу консервативних транспозонів. Він не копіюється, а переміщується з одного місця в інше за принципом "вирізати-вставити".Характеристики:Tn3 містить гени, що кодують транспозазу і бета-лактамазу (фермент, що забезпечує стійкість до антибіотиків, таких як пеніцилін).Включає інвертовані кінцеві повтори для розпізнавання транспозази.Механізм:Tn3 вирізається з одного ділянки бактеріальної ДНК і вставляється в іншу. Цей механізм переміщення сприяє поширенню генів стійкості до антибіотиків серед бактерій.Приклади:Зустрічається у багатьох патогенних бактерій, де він переносить гени стійкості до антибіотиків.
Автономні
Автономні транспозони є мобільними елементами, які містять усі необхідні гени для самостійного переміщення. Вони можуть копіювати або вирізати себе і вставляти в нові ділянки геному без зовнішньої допомоги.Структура:Містять гени, які кодують транспозазу або зворотну транскриптазу (для ретротранспозонів).Інвертовані або LTR-послідовності на кінцях.Механізм:Самостійно здійснюють транспозицію, оскільки містять усі необхідні гени для мобільності.

Ac елементи в кукурудзі
Ac (Activator) — це автономний транспозон, що здатний самостійно переміщуватися в геномі рослин, зокрема в кукурудзі. Він був відкритий Барбарою МакКлінток у 1940-х роках і є частиною її досліджень рухомих генетичних елементів.Характеристики:Ac має гени, що кодують транспозазу, яка необхідна для переміщення.Він може ініціювати як власну транспозицію, так і транспозицію неавтономних елементів, таких як Ds.Механізм:Ac вирізається з однієї частини геному і вставляється в іншу. Це відбувається за допомогою транспозази, яку він кодує, і відбувається за механізмом "вирізати-вставити".Приклади:Зустрічається в кукурудзі, де він може впливати на колір зерна через його здатність до переміщення і впливу на експресію генів.
Неавтономні
Неавтономні транспозони не можуть самостійно переміщатися через відсутність генів, що кодують білки для транспозиції. Вони потребують допомоги автономних транспозонів, щоб переміщатися в геномі.Структура:Відсутність генів для транспозиції (транспозази або зворотної транскриптази).Мають інвертовані повтори або інші послідовності, що можуть розпізнаватися білками автономних транспозонів.Механізм:Переміщення можливе лише за наявності автономних транспозонів, які надають необхідні білки для транспозиції.

Ds елементи в кукурудзі
Ds (Dissociation) — це неавтономний транспозон, який залежить від Ac для переміщення. Ds не містить гена для транспозази, тому не може самостійно переміщуватися і потребує наявності Ac.Характеристики:Відсутність власного гена транспозази робить Ds залежним від активності Ac.Ds елементи часто спричиняють мутації через переміщення в функціональні ділянки геному, впливаючи на гени.Механізм:Ds може переміщуватися тільки тоді, коли Ac виробляє транспозазу. Це також відбувається за механізмом "вирізати-вставити".Приклади:Як і Ac, Ds зустрічається в кукурудзі і був частиною відкриттів Барбари МакКлінток. Він впливає на мутації і може змінювати експресію генів, зокрема генів, відповідальних за колір зерен кукурудзи.

У еукаріотів
У геномах еукаріотів транспозони можуть становити значну частину ДНК. Наприклад:У геномі людини ретротранспозони складають майже 45% ДНК. Серед них найпоширенішими є Alu-повтори, що відносяться до SINE-ів.У рослин, таких як кукурудза, транспозони також займають великий відсоток геному та відіграють важливу роль у формуванні генетичної варіативності.

Роль в онкогенезі
Транспозони можуть відігравати важливу роль у розвитку ракових захворювань (онкогенезі), оскільки їх переміщення може призводити до порушень нормальної роботи генів:Інактивація генів-супресорів пухлин. Якщо транспозон вставляється у ген, який відповідає за контроль клітинного поділу, це може призвести до неконтрольованого зростання клітин і розвитку раку.Активація онкогенів. Транспозони можуть активувати онкогени, які за нормальних умов не експресуються. Це також може стимулювати розвиток раку.Рекомбінаційні події. Переміщення транспозонів може призводити до хромосомних перебудов (делецій, дуплікацій), які пов’язані з онкогенезом.Порушення генетичної стабільності. Мобільність транспозонів може створювати хаос у геномі, порушуючи генетичну стабільність, що є однією з характеристик ракових клітин.

Мітохондріальний геном
Кожна людина в своєму клітинному ядрі має приблизно 5 послідовностей ДНК, котрих мати б не повинна. Адже ці фрагменти ДНК, що завдовжки до 1500 пар основ, походять із мітохондрій, засвідчили аналізи. Тобто частини мітохондріального ДНК проникають у геном клітинного ядра – і цей процес триває донині. Це суперечить поширеній теорії та кидає нове світло на еволюцію нашого геному.

Особливості ДНК
Мітохондріальна ДНК або мтДНК — кільцева молекула ДНК, локалізована в мітохондріях, цитоплазматичних органелах більшості клітин еукаріотів, що мають вигляд ниткоподібних або гранулярних утворень. Локалізація мтДНК відрізняється від локалізації більшості ДНК еукаріотів, розташованої в ядрах клітин. Часто стверджується, що мітохондріальна ДНК успадковується тільки по материнській лінії, але 2018 року були знайдені 17 осіб із 3 сімей, які успадкували мтДНК від обох батьків.Ілюстрація розташування мітохондріальної ДНК у клітинах людини стала першою значною частиною людського геному, що вдалося секвенувати. Це секвенування показало, що мтДНК людини включає 16569 пар основ і кодує 13 білків.
a
Генетичний дрейф
Генетичний дрейф відбувається тому, що більш-менш копії алеля випадково можуть передаватися наступному поколінню. Це може статися тому, що випадково особини, що несуть певний алель, можуть залишити більш-менш потомство в наступному поколінні. У статевій популяції генетичний дрейф відбувається також тому, що передача Менделя означає, що потомству передається лише один з двох алелів у особини, обраний випадковим чином у місці.Генетичний дрейф може відігравати певну роль у динаміці всіх алелів у всіх популяціях, але він відіграє найбільшу роль для нейтральних алелів.Генетичний дрейф, за відсутності нових мутацій, повільно очищає наше населення від нейтрального генетичного різноманіття, оскільки алелі повільно дрейфують на високі або низькі частоти і з часом втрачаються або фіксуються.
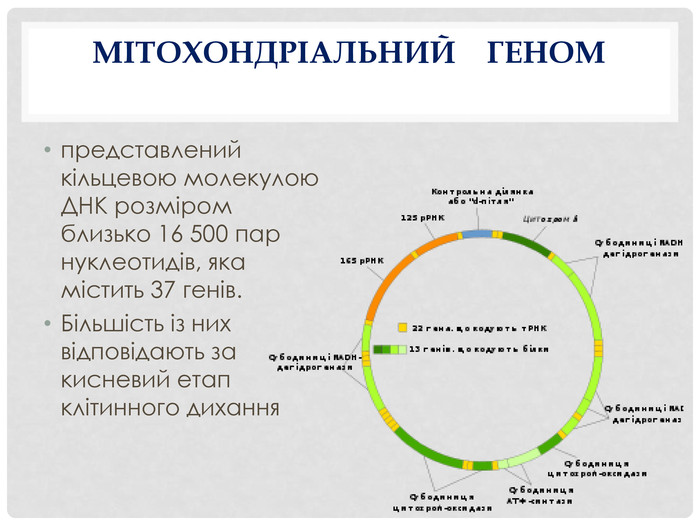
Мітохондріальний геном
Це сукупність усієї генетичної інформації, що міститься в мітохондріях. Тобто це і є вся мітохондріальна ДНК.Терміни "геном" і "ДНК" іноді використовуються з деякою різницею в контекстах, де "геном" акцентує увагу на всій сукупності генів і генетичній структурі, а "ДНК" може акцентувати на хімічній основі (молекулі ДНК).
Порівняння з ядерним геномом
Розмір геному:Мітохондріальний геном (мтДНК) значно менший за ядерний. У людини він складається приблизно з 16 569 пар нуклеотидів і містить 37 генів.Ядерний геном (ДНК, що знаходиться в ядрі клітини) набагато більший — він містить близько 3 мільярдів пар основ і включає приблизно 20 000–25 000 генів.Структура ДНК:Мітохондріальна ДНК зазвичай має кільцеву форму і існує у вигляді кільцевої молекули в мітохондріях.Ядерна ДНК є лінійною і організована у вигляді хромосом (у людини 23 пари хромосом).Походження та спадковість:Мітохондріальна ДНК передається по материнській лінії. Всі мітохондрії дитини походять від матері, оскільки яйцеклітина надає мітохондрії під час запліднення.Ядерна ДНК спадкується від обох батьків. Половина хромосом походить від батька, а половина — від матері.Кількість копій:Мітохондріальна ДНК наявна в багатьох копіях у кожній мітохондрії, а клітини містять сотні або навіть тисячі мітохондрій, що призводить до наявності багатьох копій мтДНК у кожній клітині.Ядерна ДНК представлена лише двома копіями кожної хромосоми в диплоїдних клітинах (одна копія від матері, інша від батька).Генетичний код:Мітохондріальний геном має деякі відмінності в генетичному коді порівняно з ядерним. У деяких випадках ті ж самі кодони, які кодують амінокислоти в ядерній ДНК, мають інші значення в мітохондріальній ДНК.Ядерний геном використовує стандартний генетичний код.Регуляція та контроль:Мітохондріальна ДНК менш контрольована складними регуляторними механізмами, характерними для ядерної ДНК. Вона має незначну кількість некодуючих послідовностей (інтронів майже немає), а велика частина генів безпосередньо кодує білки, РНК або інші важливі компоненти.Ядерна ДНК має багато складних регуляторних елементів і містить значну частину некодуючих послідовностей (інтрони, промотори, енхансери тощо).Функції генів:Гени в мітохондріальному геномі головним чином відповідають за виробництво енергії (через дихальний ланцюг і синтез АТФ) і синтез білків, що потрібні для роботи мітохондрій.Ядерний геном регулює більшість процесів у клітині, включаючи структурні білки, ферменти, клітинний цикл, сигнальні шляхи та всі інші біохімічні процеси, які відбуваються в організмі.
Еукаріоти
Еукаріоти – організми (вищі тварини і рослини, гриби, одно- і багатоклітинні водорості), клітини яких містять сформоване ядро; ядерна ДНК з'єднана з гістонами і негістоновими білками і організована у вигляді хроматину/ хромосом. Термін запропонований Е. Шаттоном в 1937 р.

Поліплоїдія
Поліплоїдія — збільшення кількості наборів хромосом в клітках тканин організму, кратне одинарному (гаплоїдному) набору хромосом – молекулярних структур, де міститься близько 90 % ДНК клітини.Поліплоїди – це організми, в клітинах яких є більше двох наборів хромосом: триплоїди – містять три набори, тетраплоїди – містять чотири набори і т.д. Поліплоїди з непарним набором хромосом (3n, 5 n) стерильні, тобто вони не діляться і як результат – не дають потомства. Яскравим прикладом поліплоїду з непарним набором хромосом є банани, які не розмножуються насінням, а лише вегетативним шляхом.
a
Автополіплоїдія
Автополіплоїдія — більше ніж дворазове спадкове кратне збільшення (зазвичай, спонтанне) ідентичних наборів хромосом у клітинах рослин, рідше тварин, того самого виду. Також виникнення поліплоїдного організму внаслідок геномної иутації типу поліплоїдії у предкового диплоїдного організму аналогічного виду.
a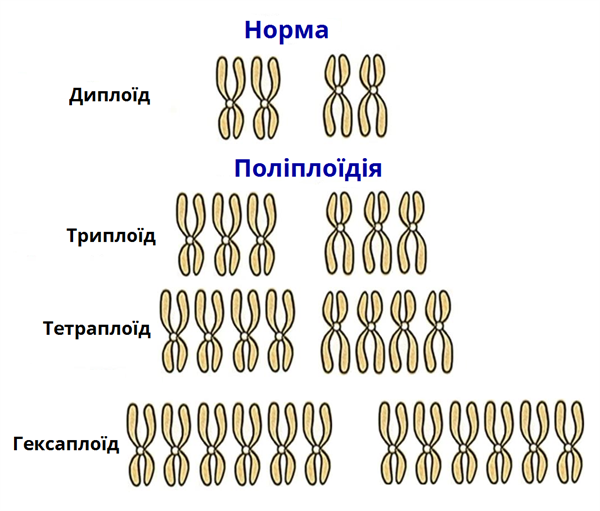
Алополіплоїдія
Поєднання в клітинах організмів наборів хромосом - геномів, що походять від різних видів або родів, причому один, два чи кожний з геномів представлений більше як один раз. А. являє собою поліплоїдію на основі гібридизації. Алополіплоїди, що поєднують по 2 геноми різних видів, наз. амфідиплоїдами, або алотетраплоїдами, 2 геноми одного виду і 1 другого - сескві-плоїдами. А. є одним з головних шляхів подолання безплідності міжвидових гібридів рослин; має значення для видоутворення; використовується в селекції рослин.
a
Значення для еволюції
Варіативність: Поліплоїдія може створювати нові генетичні варіації, які можуть бути корисними в процесі еволюції. Це надає організмам можливість пристосовуватися до нових умов середовища.Генетична стабільність: Поліплоїдні рослини часто є більш життєздатними та стійкими до стресів (наприклад, посухи, хвороб).Економічне значення: Багато сільськогосподарських культур є поліплоїдними, що робить їх продуктивнішими та більш придатними для вирощування.
aЛокалізація геному
Невеликі ділянки ДНК, які називаються генами, і складаються з певних послідовностей нуклеотидів, кодують молекули РНК, з інформації яких синтезуються білкові молекули, необхідні організму. В еукаріотів геном кожної клітини міститься в клітинному ядрі, оточеному мембраною.
Ядро
Клітинне ядро — обов'язковий компонент будь-якої еукаріотичної клітини і є її найважливішою частиною.Від цитоплазми ядро відокремлене оболонкою, що складається з двох мембран. В оболонці ядра є численні пори, які потрібні для того, щоб різні речовини могли потрапляти з цитоплазми в ядро і навпаки.Існує два типи ядер:генеративні — забезпечують зберігання і передачу спадкової інформації; вегетативні — регулюють біосинтез білка.
Мітохондрії
Мітохондрії – це крихітні субодиниці присутні у кожній клітині людського тіла від шкіри до внутрішніх органів, крім еритроцитів. Основна роль мітохондрій полягає у тому, щоб перетворити їжу та кисень на корисну енергію – АТФ, необхідну організму для підтримки життєдіяльності. Відомо також, що ці органели беруть активну участь у біосинтетичних процесах, пов’язаних із поліпептидним, амінокислотним та жировим катаболізмом, формуванням та метаболізмом сечовини, органічних кислот, у біосинтезі гему, нуклеотидів, стероїдів, кардіоліпіну, убіхінону, різних метаболітів та сигнальних молекул.
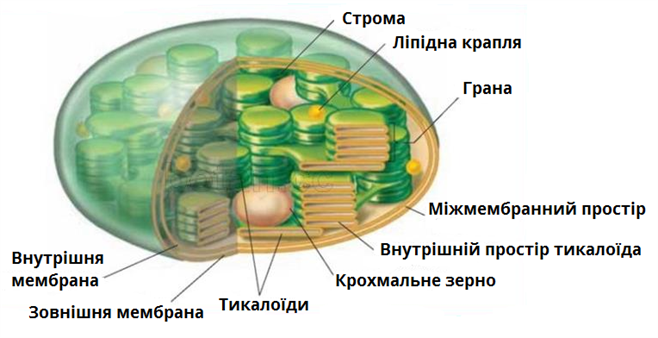
Хлоропласти
Хлоропласти - це великі органели і їх функція полягає в утворенні і зберіганні вуглеводів з фотосинтезу. Хлоропласт обмежений подвійною мембраною.Матрикс хлоропласту відомий як строма. Також всередині хлоропласта знаходяться окремі внутрішні мембрани, які утворюють ламелі або округлі тилакоїди, схожі на язик всередині огороджувальної подвійної мембрани. Ці язикоподібні або дископодібні тилакоїдні мембрани можуть бути складені шарами, і вони називаються гранами. Грани з'єднуються між собою за допомогою інших мембран.
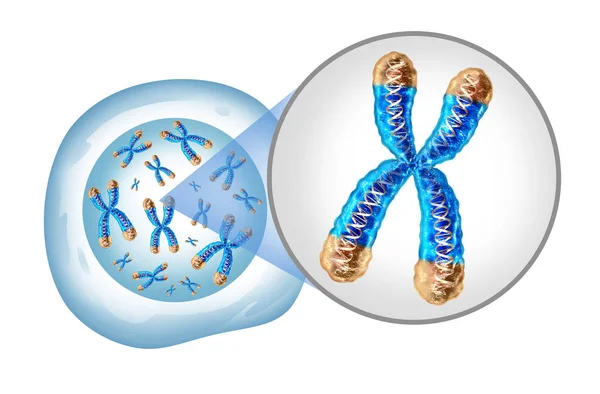
Хромосоми і їх структура
Хромосоми (грец. сhróma — колір, забарвлення + sóma — тіло) — основні структури клітинного ядра, які є носіями генів і визначають спадкові властивості клітин та організмів. Здатні до самовідновлення, їм притаманна структурова та функціональна індивідуальність, вони зберігають її в ряді поколінь. Термін Х. запропонований В. Вальдейєром (1888). Основу Х. складає одна безперервна дволанцюгова молекула ДНК (близько 99% ДНК клітини міститься в Х. ядра), зв’язана з білками (гістонами та ін.) в нуклеопротеїновий комплекс. Будовою молекул ДНК, її генетичним кодом забезпечується запис спадкової інформації в Х., білки (в Х. вищих рослин та тварин їх міститься до 65%) беруть участь у складному упакуванні ДНК в Х. і регуляції її здатності до синтезу РНК — транскрипції. У процесі функціонування Х. підлягають структурно-морфологічним перетворенням, в основі яких лежить процес спіралізації-деспіралізації структурових субодиниць Х. — хромонем (хромосомний цикл). На стадії метафази поділу клітин (див. Мітоз, Мейоз) спіралізовані (щільно упаковані) Х. добре розрізняються в світловому мікроскопі. Кожна Х. складається з двох поздовжніх копій — хроматид, які утворилися в ході редуплікації і скріплені центромерою. У клітинах тіла двостатевих тварин і рослин кожна Х. представлена двома так званими гомологічними Х., одна з яких походить від материнського організму, а інша — від батьківського.
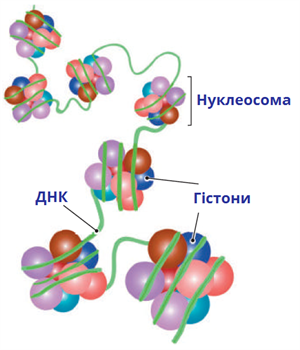
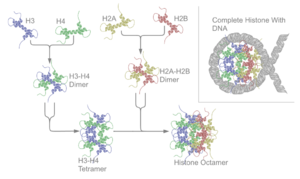
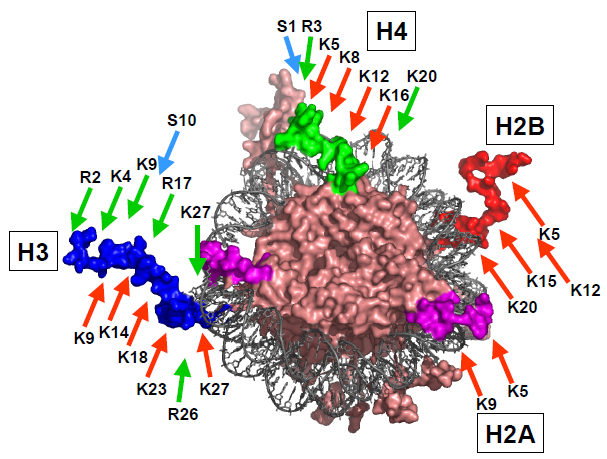
Гістони
Гістони (грец. histos — тканина) — група висококонсервативних простих білків, асоційованих з ДНК та негістоновими білками у складі хроматину ядра клітини. Г. відкриті у 1884 р. німецьким біохіміком А. Косселем. Мол. м. Г. у середньому становить 10 000 — 23 000 Да. Г. мають виражені основні властивості. Для первинної структури Г. характерна велика кількість основних амінокислот — лізину та аргініну, які переважно знаходяться на N- і C-кінцевих ділянках молекули білка. Крім того, Г. містять мало цистеїну, метіоніну і практично не містять триптофану. Внутрішні ділянки молекул Г. є надзвичайно консервативними в еволюційному плані та містять переважно гідрофобні амінокислоти. Залежно від розміру молекули та вмісту окремих амінокислот Г. поділяють на 5 класів: Н1, Н2А, Н2В, Н3, Н4. Г. є головним білковим компонентом хромосом. Основна біологічна роль Г. — участь у стабілізації просторової структури ДНК та утворенні нуклеопротеїнових комплексів — нуклеосом, які становлять глобули і є головними структурними одиницями хромосом. Білкове ядро нуклеосоми, або нуклеосомний кор, складається з октету Г.: тетрамеру (Н3)2-(Н4)2 та двох димерів Н2А-Н2В, які зв’язуються один з одним за допомогою гідрофобних взаємодій. На поверхню нуклеосомного кору намотується відрізок ДНК завд. у 140–160 нуклеотидних пар, утворюючи лівозакручену суперспіраль.

H1
H1 є лінкерним гістоном, який не входить до складу нуклеосом, а допомагає з'єднувати нуклеосоми між собою.Функція: Підтримує структурну організацію хроматину, сприяє формуванню більш компактних структур.
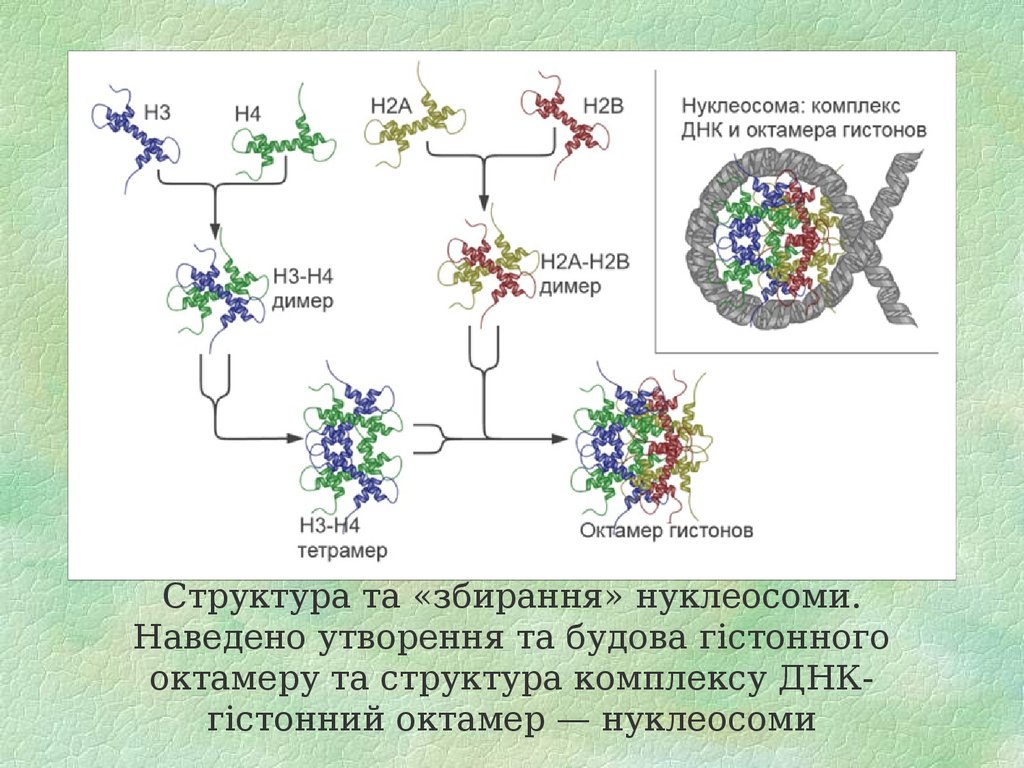
H2A
Один з основних гістонів, який входить до складу нуклеосоми.Функція: Сприяє упаковці ДНК та формуванню нуклеосом. Існує кілька варіантів H2A, включаючи H2A.Z, який грає роль у регуляції генів.
H2B
Ще один основний гістон, який входить до складу нуклеосом.Функція: Допомагає в організації та стабілізації нуклеосом, впливаючи на доступність ДНК для транскрипції та реплікації.
H3
Третій основний гістон, який входить до складу нуклеосом.Функція: Важливий для організації хроматину. Модифікації H3, такі як метилювання та ацетилювання, можуть впливати на експресію генів.
H4
Останній з основних гістонів, який також входить до складу нуклеосом.Функція: Спільно з H2A, H2B та H3 утворює ядерну структуру (нуклеосому), важливу для упаковки ДНК.
Модифікація
Модифікації змінюють заряд, гідрофобність та інші властивості поверхні глобул. Це може призводити до змін у просторовій структурі ДНК. Модифікації Г. є одним із механізмів регуляції передачі спадкової інформації, зокрема процесів реплікації і транскрипції, у клітинах еукаріотів

Метилювання
У дуже спрощених термінах можна сказати, що метилювання функціонує як перемикач: увімкнення / вимкнення у багатьох контекстах. І гени, і певні молекули можуть бути активовані або дезактивовані шляхом метилювання. Він важливий у багатьох реакціях трансформації в організмі і, отже, забезпечує ряд важливих речовин.Але що саме таке метилювання? Метилювання — а точніше трансметилювання — означає просто перенесення метильних груп від однієї молекули до іншої. Опосередкована певними ферментами, одна молекула вивільняє метильну групу, а інша поглинає її. Ця проста реакція часто забезпечує активацію певних речовин і безліч метаболічних процесів.Щоб відбулося метилювання, організм залежить від цілого ряду ферментів, які забезпечують перенесення метильних груп між певними молекулами. Не лише один фермент сприяє метилюванню; багато різних реакцій метилювання між речовинами потребують власного ферменту.
Репресія генів
Блокування експресії генів називається репресією. Білки-репресори кодуються генами, які називають регуляторними. Репресори зв'язуються з ділянками ДНК на початку гену, операторами, у такий спосіб запобігаючи зв'язуванню з ним РНК-полімерази та синтезу нею мРНК.
Активація генів
Активація генів — це процес, за допомогою якого ген або його експресія стають активними, що призводить до синтезу відповідного білка або РНК. Цей процес включає ряд молекулярних подій, які перетворюють інформацію, закодовану в ДНК, на функціональні молекули. Активація генів може відбуватися через кілька механізмів, таких як:Модифікації гістонів: Наприклад, ацетилювання гістонів зменшує їх взаємодію з ДНК, що дозволяє більш легкому доступу до генетичної інформації.Зв'язування регуляторних білків: Специфічні транскрипційні фактори можуть зв'язуватися з промоторами або посилювачами генів, стимулюючи їх активність.Репрограмування хроматину: Зміни в структурі хроматину, такі як відкриття або закриття хроматинового комплексу, також можуть активувати гени.Сигнальні шляхи: Різноманітні сигнальні молекули (гормони, фактори росту) можуть активувати або пригнічувати експресію генів, впливаючи на відповідні механізми на рівні клітини.
Ацетилювання
Ацетилювання — це процес, при якому ацетильна група (—COCH₃) прикріплюється до аміногрупи лізину. Це є однією з найпоширеніших модифікацій гістонів.Класифікація:Ацетилювання: Зазвичай відбувається на лізинах (наприклад, H3K9ac, H3K27ac).Функції:Активація генів: Ацетилювання гістонів зазвичай асоціюється з активними генами. Ацетильна група нейтралізує позитивний заряд лізину, що зменшує взаємодію з негативно зарядженим фосфатом ДНК, і таким чином сприяє розслабленню хроматину.Взаємодія з регуляторами: Модифікації ацетилювання створюють місця для зв'язування інших білків, які відповідають за активацію гена.

Фосфорилювання
Фосфорилювання — це процес приєднання фосфатної групи (—PO₄) до гістонів, зазвичай на серинових або треонінових залишках.Класифікація:Фосфорилювання на серині: Наприклад, H3S10ph.Фосфорилювання на треоніні: Наприклад, H3T3ph.Функції:Регуляція клітинного циклу: Фосфорилювання гістонів часто асоціюється з переходом між фазами клітинного циклу, такими як поділ клітини.Активація або репресія генів: Залежно від конкретних контекстів і модифікацій, фосфорилювання може як активувати, так і репресувати гени.Асоціація з репарацією ДНК: Фосфорилювання H2AX (гистону H2A) відбувається у відповідь на ушкодження ДНК, що сигналізує про участь у процесах репарації.
Хроматин
Хроматин (грец. сhróma — колір, забарвлення) — нуклеопротеїнові нитки, з яких складаються хромосоми клітин еукаріотів. Термін уведений В. Флемінгом (1880). У цитології під Х. розуміють дисперсний стан хромосом в інтерфазі клітинного циклу. Основні структурні компоненти Х. — ДНК (30–45%), гістони (30–50%) та негістонові білки (4–33%). На електронних мікрофотографіях Х. нагадує намисто, утворене нуклеосомами — часточками діаметром близько 11 нм. Вищі порядки структурової організації Х. (хромосоми) утворюються з лінійного пучка елементарних ниток Х. — хромомерів — за рахунок суперспіралізації, утворення петель прикріплення до «осьового скелета» з негістонових білків. У цих процесах беруть участь гістони, іони металів і т.д.
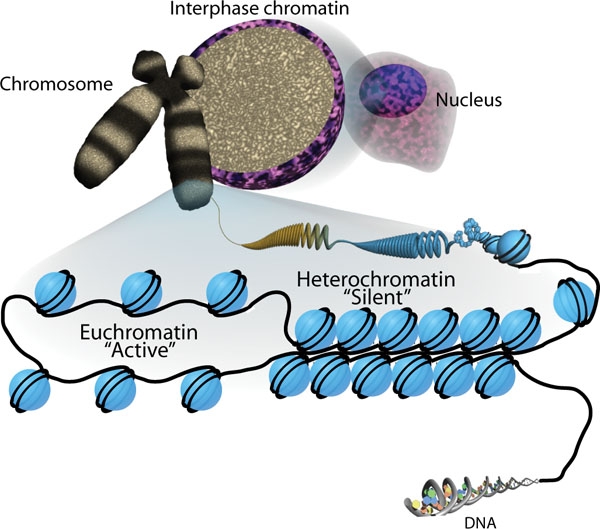
Еухроматин
Еухромати́н — стан хроматину (ДНК в ядрі) який характеризується "відкритістю" та транскрипційною активністю — з еухроматину зчитується РНК. В противагу еухроматину, гетерохроматин — стан ДНК, що щільно запакований і не є транскрипційно активним.
Гетерохромати
Гетерохроматин — конденсований стан хроматину, що утворює хромоцентри в ядрі на стадії інтерфази, а також ділянки інтенсивного забарвлення на метафазних хромосомах. Особливістю гетерохроматину є транскрипційна інертність ДНК, що входить до його складу.
Лінійні хромосоми
Лінійні хромосоми — це тип хромосом, які мають лінійну, а не кільцеву структуру. Вони складаються з довгих молекул ДНК, що містять генетичну інформацію, та специфічних білків, зокрема гістонів, які організують цю ДНК у компактну форму, щоб вона могла поміститися в клітинному ядрі.
Епігенетична регуляція
Епігенетична регуляція — це процес, за допомогою якого змінюється експресія генів без зміни послідовності ДНК. Це може відбуватися через модифікації гістонів, метилювання ДНК, а також завдяки впливу некодувальних РНК, що змінюють активність генів, впливаючи на їх доступність для транскрипції.
Метилювання ДНК
Метилювання ДНК — модифікація молекули ДНК без зміни її нуклеотидної послідовності. Основний механізм епігенетики. Метилювання ДНК полягає в приєднанні метильної групи до позицій C-5 або N-4 цитозину або позиції N-6 аденіну.Загалом метилювання впливає на рівень транскрипції, і тому є частиною регулювання експресії генів. Інформація про метилювання може наслідуватися із поділом клітини, і таким чином може розглядатися як частина епігенетичного коду, епігенетичної складової геному.
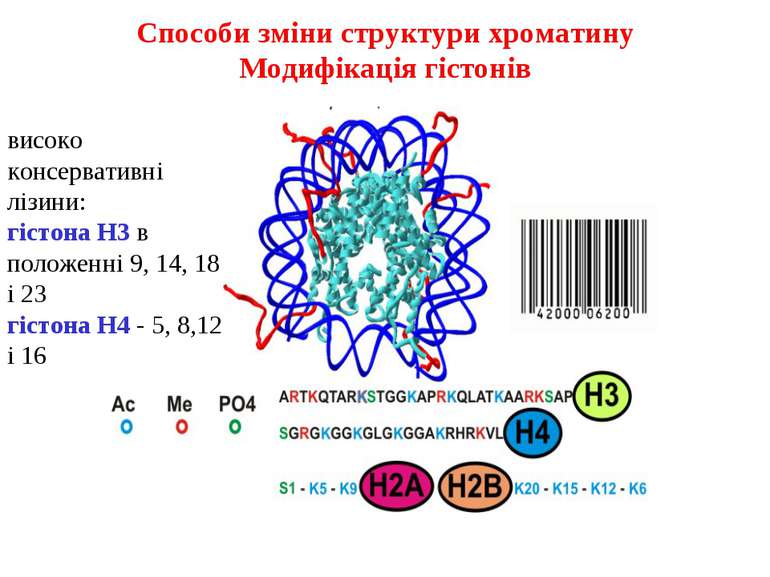
Модифікація гістонів
Модифікації гістонів - фосфорилювання, метилювання і ацетилювання призводять до зміни структури хроматину, який зумовлює зміну рівнів експресії генів. Фосфорилювання і ацетилювання носять динамічний характер - можуть досить швидко виникати і зникати. Метилювання гістонів носить найбільш статичний характер.
a
Сплайсинг
Сплайсинг РНК — процес «вирізання» інтронів і «зшивання» екзонів під час процесингу попередника матричної РНК, синтезованого в результаті транскрипції; також існує сплайсинг тРНК. Через те, що в геномах прокаріотів інтрони дуже рідкі, сплайсинг зазвичай характерний для еукаріотів.
Альтернативний сплайсинг
Альтернативний сплайсинг — механізм виникнення варіацій при сплайсингу РНК за рахунок відокремлення екзонів попередника мРНК (пре-мРНК) і їх повторного з'єднання. В результаті отримуються альтернативні варіанти нуклеотидної послідовності мРНК. Ці варіанти мРНК, мРНК-ізоформи. або сплайс-варіанти, потім зазвичай використовуються у процесі трансляції та переводяться в амінокислотну послідовність, формуючи ізоформи білків. Таким чином, альтернативний сплайсинг збільшує різноманітність білків, що синтезуються клітиною.

Стоп кодони
Стоп-кодон визначає закінчення (термінацію) синтезу поліпептидного ланцюга. Це кодони УАА, УАГ, УГА. Термінуючими кодонами можуть бути також кодони АГА й АГТ; натомість, кодон УГА в мРНК, що транскрибується з мітохондріального генома (крім вищих рослин), не є термінуючим, а кодує триптофан.
Точки сплайсингу
Точка сплайсингу — це специфічна ділянка на РНК, де відбувається процес вирізання інтронів і з'єднання екзонів під час дозрівання пре-мРНК у зрілу мРНК. Цей процес забезпечує правильне формування білків відповідно до генетичної інформації.
Виключення екзонів
Транс-сплайсинг
Транс-сплайсинг - форма сплайсингу, при якій з'єднуються РНК різних транскриптів. У трипаносом, нематод і деяких інших нижчих еукаріотів описані випадки транс-сплайсингу, в результаті якого до мРНК пришивається коротка лідерна послідовність, яка приходить з іншої частини геному.
Автосплайсинг
Автосплайсинг — це процес самостійного вирізання інтронів і з'єднання екзонів у молекулі РНК без участі сплайсосом або інших білкових комплексів. Цей механізм спостерігається в деяких рибозимах — молекулах РНК, які мають ферментативну активність і можуть каталізувати власне сплайсування.

Прокаріоти
Прокаріоти – організми, клітини яких позбавлені обмеженого мембраною ядра (аналогом ядра є нуклеоїд). Зараз прокаріоти найчастіше поділяють на два домени: еубактерій і архей. Клітини прокаріотів позбавлені більшості мембранних (хлоропластів, мітохондрій, апарату Гольджі) і утворених мікротрубочками органел (центріолей, джгутиків і війок з мікротрубочок) еукаріотичної клітини, а рибосоми значно відрізняються від рибосом еукаріотів. Термін запропонований в 1937 р. Е. Шаттоном.

Мутації
Деякі мутації позитивно впливають на організм, в якому вони відбуваються. Їх відносять до корисних мутацій. Вони, як правило, кодують нові версії білків, які допомагають організмам адаптуватися до навколишнього середовища. Якщо вони збільшують шанси організму на виживання або розмноження, мутації, швидше за все, з часом стануть більш поширеними. Існує кілька відомих прикладів корисних мутацій. У бактерій відбулися мутації, які дозволяють бактеріям виживати в присутності антибіотичних препаратів. Мутації призвели до еволюції стійких до антибіотиків штамів бактерій.
Виникнення
Мутації як еволюційний механізм: у прокаріотів мутації є одним із ключових механізмів еволюції через їхню високу швидкість поділу клітин. Це дозволяє бактеріям швидко адаптуватися до нових середовищ або змінювати механізми стійкості до антибіотиків.
a
Швидкість мутацій
Швидкість мутацій у прокаріотів може варіювати в залежності від виду і умов. У середньому для бактерій вона складає близько 1 мутації на кожен 10⁶-10⁹ поділів клітин. У деяких випадках мутації можуть відбуватися швидше:"Гіпермутації" — деякі штами бактерій мають високу частоту мутацій через пошкодження механізмів репарації ДНК.Фактор стресу — у стресових умовах (наприклад, нестача поживних речовин або дія антибіотиків) деякі бактерії можуть збільшувати частоту мутацій, що сприяє швидшому пристосуванню.
Наслідки мутацій для бактерій
Мутації можуть мати різноманітні наслідки для прокаріотів:Шкідливі мутації можуть призвести до порушення функцій важливих генів, що знижує життєздатність або вбиває клітину.Нейтральні мутації не мають помітного впливу на функціонування клітини.Корисні мутації можуть покращувати здатність бактерій до виживання в певних умовах, наприклад, забезпечувати стійкість до антибіотиків або підвищувати ефективність утилізації нових джерел енергії.
a
Плазміди
Деякі характеристики плазмід роблять їх ідеальними векторами (транспортними засобами для перенесення послідовностей ДНК від одного організму до іншого) для генної інженерії, наприклад:Здатність реплікуватися, тобто робити свої копії незалежно від бактеріальної хромосоми. Для цього плазміди мають специфічну послідовність, де ферменти синтезу ДНК клітини-хазяїна зв’язуються та ініціюють реплікацію ДНК (біологічний процес, який відбувається в усіх живих організмах для створення копій їх ДНК). Ця послідовність називається сайтом ori («місце початку реплікації» ).Здатність ініціювати транскрипцію (процес, за допомогою якого інформація, закодована в ДНК, передається на інформаційну РНК за допомогою РНК-полімеразиклітини-господаря). Для цієї здатності потрібна інша специфічна послідовність, яка називається промоторною послідовністю. Промоторна послідовність з'єднується з РНК-полімеразою; тут починається транскрипція. Усі гени мають промоторні послідовності, розташовані поруч з ними в ДНК. Для того щоб гени, такі як ген інсуліну, могли експресуватися в бактеріях, вони повинні бути вставлені в плазміду поряд з промоторною послідовністю.Ген або гени, що кодують стійкість до антибіотиків, класу сполук, які вбивають або пригнічують ріст мікроорганізмів. Ці гени кодують білки, що виділяються мікроорганізмами, які пригнічують дію антибіотиків, і можуть надавати селективну перевагу в природі бактеріям, що містять цю плазміду, у мікробній популяції, в якій бактерії змагаються за виживання.

F-плазміди
F-плазміди, або фертильні плазміди, є особливим класом плазмід, які несуть гени, що відповідають за процес кон'югації – обмін генетичним матеріалом між бактеріями.Характеристики:Розмір: F-плазміди можуть бути досить великими, зазвичай мають розмір близько 100 тисяч пар основ (100 kbp).Наявність генів кон'югації: Містять гени, які кодують білки для утворення пілі — спеціалізованої структури, через яку бактерії можуть передавати плазміду іншій клітині.Фактор F (F-фактор): Це плазміда, яка здатна переносити копію себе через кон'югаційний міст між бактеріями, що створює можливість для обміну генетичним матеріалом між різними клітинами. Бактерії з F-фактором називають F+ (ті, що мають плазміду), а без нього — F- (ті, що не мають плазміду).
R-плазміди
R-плазміди — це плазміди, що містять гени стійкості до антибіотиків, і є одним з головних факторів поширення антибіотикорезистентності серед бактерій.Характеристики:Розмір: R-плазміди можуть бути різних розмірів, від кількох до сотень тисяч пар основ.Гени резистентності: R-плазміди несуть гени, які кодують білки, що забезпечують стійкість до різних антибіотиків. Це можуть бути гени, що відповідають за модифікацію антибіотиків, їхню деградацію або ефлюкс (виведення з клітини).

R100 плазміда
R100 плазміда — це добре вивчена плазміда резистентності (R-плазміда), яка забезпечує бактеріям стійкість до кількох різних антибіотиків. Вона є однією з найвідоміших плазмід, які відіграють важливу роль у поширенні антибіотикорезистентності серед патогенних бактерій.Основні характеристики R100 плазміди:Розмір: Плазміда R100 має приблизно 94-100 тисяч пар основ (94-100 kbp).Гени резистентності:R100 плазміда містить гени, що забезпечують стійкість до кількох різних класів антибіотиків. Вона кодує ферменти або білки, які нейтралізують дію антибіотиків або перешкоджають їхньому потраплянню в клітину.Гени, що кодують стійкість до таких антибіотиків, як:ТетрациклінСульфаніламідиХлорамфеніколСтрептоміцинКанаміцинМеханізм дії:Експорт антибіотиків: Деякі гени, присутні в плазміді R100, кодують білки, що діють як ефлюксні насоси. Вони виводять антибіотики з клітини, знижуючи їх концентрацію до неефективного рівня.Модифікація антибіотиків: Інші гени кодують ферменти, які хімічно модифікують антибіотики, роблячи їх неактивними.Адаптація клітинних процесів: Деякі з генів дозволяють бактеріям уникати цільової дії антибіотиків, змінюючи свої внутрішньоклітинні процеси.Мобільність:R100 плазміда належить до кон'югативних плазмід, що означає, що вона може передаватися від однієї бактерії до іншої через процес кон'югації. Вона містить гени, які дозволяють утворювати пілі для передачі генетичного матеріалу.Транспозони:В плазміді R100 можуть бути інтегровані транспозони, які забезпечують додаткову стійкість до антибіотиків та дозволяють плазміді швидше поширюватися між різними бактеріями.Клінічне значення:R100 плазміда становить серйозну проблему для лікування інфекцій, викликаних патогенними бактеріями, оскільки забезпечує стійкість до багатьох антибіотиків. Плазміди такого типу можуть передаватися між патогенами, такими як Escherichia coli і Salmonella, що робить їх важливими елементами в поширенні антибіотикорезистентності.
aSalmonella
Сальмонела — рід грам-негативних бактерій, родини ентеробактерій, що спричинюють черевний тиф, паратифи і сальмонельози. Види Salmonella рухомі за допомогою джгутиків і виробляють сірководень.
a
Escherichia coli
Кишкова́ паличка — відкрита німецьким педіатром і мікробіологом Теодором Ешеріхом — один з головних видів бактерій, що живуть у нижніх відділах кишечнику ссавців, загалом відомих як флора кишечника. Деякі зразки також були знайдені на краю гарячих джерел.
Резистентність до антибіотиків
Основною функцією R-плазмід є надання бактеріям здатності протистояти дії антибіотиків, таких як тетрациклін, хлорамфенікол, ампіцилін і інші.
a
Швидке поширення резистентності
R-плазміди можуть передаватися від однієї бактерії до іншої через кон'югацію, що сприяє швидкому поширенню антибіотикорезистентних бактерій у популяції.
Кільцева хромосома
Кільцева хромосома — це аномальний тип хромосоми, яка має кільцеву або замкнену структуру, що утворюється внаслідок злиття кінців хромосоми. У нормальних умовах хромосоми мають лінійну структуру, однак за певних обставин кінці можуть з'єднуватися, утворюючи кільце. Кільцеві хромосоми можуть утворюватися як у людей, так і у тварин та рослин.Характеристика:Утворення:Кільцеві хромосоми зазвичай формуються через втрату теломерів — кінцевих ділянок хромосоми, які забезпечують захист ДНК. Після втрати теломер кінці хромосоми можуть злипатися і формувати кільце.Інший механізм утворення включає розриви хромосоми та її подальше злиття в кільце.Механізми виникнення:Спонтанні мутації: Кільцеві хромосоми можуть виникати внаслідок спонтанних мутацій або аномалій під час поділу клітин.Екологічні фактори: Радіація, хімічні мутагени або інші стресові фактори можуть викликати розриви ДНК, що призводять до утворення кільцевих хромосом.Розповсюдження:Кільцеві хромосоми зустрічаються рідко в природі, проте відомі випадки як у людей, так і у тварин. Наприклад, у людини кільцеві хромосоми можуть утворюватися внаслідок генетичних захворювань або аномалій.
Реплікація ДНК
Реплікація, редуплікація, ауторепродукція, аутосинтез — процес самовідтворення нуклеїнових кислот, генів та хромосом. Термін застосовують переважно для опису процесу подвоєння нуклеїнових кислот, відтворення хромосом частіше описують іншими термінами.
Суперспіралізація ДНК
Суперспіралізація — це процес додаткового скручування подвійної спіралі ДНК, що дозволяє компактніше організувати її в обмеженому просторі клітини. У природі суперспіралізація ДНК зустрічається як у прокаріотичних, так і у еукаріотичних організмів.Типи суперспіралізації:Негативна суперспіралізація: Подвійна спіраль ДНК закручується у протилежному напрямку до основного напрямку спіралі. Такий тип спіралізації сприяє розкручуванню ДНК, що необхідно для процесів реплікації та транскрипції.Позитивна суперспіралізація: Спіраль закручується в тому ж напрямку, що й основна структура подвійної спіралі, що ускладнює її розкручування.Біологічне значення:Компактизація ДНК: Суперспіралізація дозволяє зменшити розмір ДНК та забезпечити її упаковку в клітину. Це особливо важливо для прокаріот, де ДНК має форму кільцевої молекули.Реплікація та транскрипція: Негативна суперспіралізація полегшує доступ до ДНК для синтезу РНК і ДНК, оскільки розкручування подвійної спіралі стає легшим.Регуляція напруги: Під час транскрипції або реплікації ДНК піддається напрузі через розкручування спіралі, і суперспіралізація допомагає компенсувати цю напругу.
Генетичні острови
Генетичні острови — це великі сегменти ДНК, які були отримані бактеріями через горизонтальний генний перенос і інтегровані в їх геном. Вони часто містять гени, які забезпечують додаткові можливості для бактерій, такі як антибіотикорезистентність, вірулентність або адаптація до нових умов. Генетичні острови відіграють важливу роль у еволюції бактерій, їх вірулентності та патогенності.
aПатогенність
Генетичні острови можуть робити бактерії більш патогенними, надаючи їм здатність виживати в організмі хазяїна або протистояти захисним механізмам організму. Наприклад, вони можуть містити гени, які відповідають за утворення біоплівок, що ускладнює боротьбу імунної системи з бактеріями.Патогенні бактерії, які мають генетичні острови, можуть швидше еволюціонувати та розширювати свій спектр хазяїв, збільшуючи ризик поширення захворювань.Приклади патогенності, пов’язаної з генетичними островами:Острів патогенності SPI-1 та SPI-2 у Salmonella: Ці острови містять гени, які забезпечують інвазію в клітини хазяїна та захист від імунної системи.Вірулентні острови у Escherichia coli: Деякі штами E. coli мають генетичні острови, що містять гени, які сприяють утворенню токсинів або протидії імунітету.
aВірулентність
Генетичні острови часто містять вірулентні фактори, такі як гени, які забезпечують бактерії здатність заражати хазяїна або протидіяти імунній системі. Це може включати токсини, адгезини (білки, які допомагають бактерії прикріплюватися до клітин) або системи секреції, які дозволяють бактерії вводити шкідливі молекули в клітину хазяїна.Завдяки швидкому поширенню вірулентних генів між бактеріями, популяції можуть ставати більш агресивними у розвитку інфекцій.
a
Вплив на еволюцію бактерій
Генетична гнучкість: Генетичні острови сприяють швидким еволюційним змінам у бактерій, оскільки дозволяють їм отримувати корисні гени через горизонтальний генний перенос, зокрема від інших бактерій, вірусів або навіть від еукаріот.Адаптація до середовища: Завдяки генам на генетичних островах, бактерії можуть швидко адаптуватися до нових екологічних умов, таких як присутність токсинів, антибіотиків або інші стресові фактори.
aГоризонтальний перенос генів
Стабільну версію було перевірено 9 квітня 2019. Горизонта́льне перене́сення ге́нів — будь-який процес, під час якого організм або клітина передає генетичний матеріал іншому організму (клітині), який не обов'язково є його нащадком.
Кон'югація
Кон'югація — це різні форми статевого процесу , за якого клітини одноклітинних (частина видів бактерій, водоростей тварин) чи багатоклітинних(деякі гриби, нитчасті зелені водорості) організмів обмінюються спадковим матеріалом.
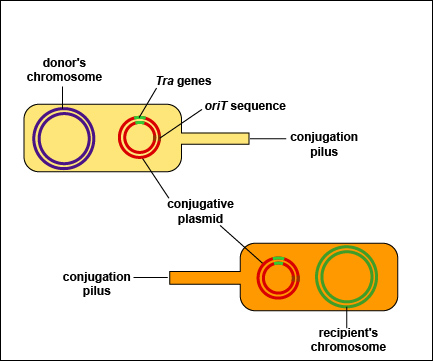
Крок 1
У грамнегативних бактерій перший крок кон'югації включає кон'югацію pilus (sex pilus або F pilus) на донорської бактерії, що зв'язується з бактерією-реципієнтом, яка не має кон'югації пілуса.
Крок 2
Зазвичай пілус кон'югації втягує або деполімеризує, витягуючи дві бактерії разом. Серія мембранних білків, закодованих кон'югативною плазмідою, потім утворює міст і отвір між двома бактеріями, тепер називається парою спаровування.
Крок 3
Використовуючи модель рухомого кола реплікації ДНК, нуклеаза розриває одну нитку плазмідної ДНК у місці походження місця перенесення (ORIT) плазміди. Нуклеаза також має хеліказовую активність і розмотує пасмо, яке буде перенесено.
Крок 4
Вирізана плазмідна нитка потрапляє в бактерію-реципієнт. Інша пасмо залишається ззаду в донорській клітині.
Крок 5
І донор, і реципієнт плазмідні пасма потім роблять додаткову копію себе.
Крок 6
Обидві бактерії тепер мають кон'югативну плазміду і можуть зробити пілус кон'югації.

Трансдукція
Трансдукція — форма горизонтального перенесення генів, при якій передача генетичного матеріалу від однієї клітини до іншої відбувається за допомогою вірусу, що, як і у випадку інших форм горизнотального перенесення генів, призводить до зміни спадкових властивостей.

Крок 1
Бактеріофаг адсорбується до чутливої бактерії.

Крок 2
Геном бактеріофага потрапляє в бактерію. Геном спрямовує метаболічний механізм бактерії на виробництво бактеріофагів компонентів та ферментів. Ферменти, кодовані бактеріофагами, також будуть розщеплювати бактеріальну хромосому.
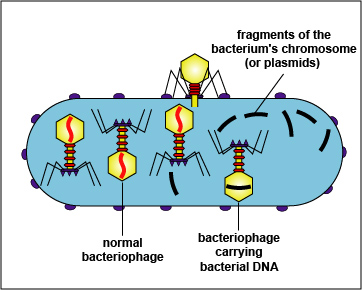
Крок 3
Іноді бактеріофаг капсид помилково збирається навколо фрагмента хромосоми донорської бактерії або навколо плазміди замість генома фага.

Крок 4
Бактеріофаги вивільняються, коли бактерія лізується. Зверніть увагу, що один бактеріофаг несе фрагмент ДНК донорської бактерії, а не геном бактеріофага.

Крок 5
Бактеріофаг, що несе ДНК донорської бактерії, адсорбується до бактерії-реципієнта.

Крок 6
Бактеріофаг вставляє ДНК донорської бактерії, яку вона переносить у бактерію-реципієнт.

Крок 7
Відбувається гомологічна рекомбінація, і ДНК донорської бактерії обмінюється на частину ДНК реципієнта.
Трансформація
Трансформація у горизонтальному генному переносі — це процес, під час якого бактерія поглинає вільну ДНК з навколишнього середовища та інтегрує її в свій геном, що дозволяє отримати нові генетичні ознаки без прямого контакту з іншими клітинами.

Крок 1
Донорська бактерія гине і деградує.

Крок 2
Фрагменти ДНК, як правило, близько 10 генів довжиною, від мертвої донорської бактерії зв'язуються з трансформасомами на клітинній стінці компетентної, живої бактерії реципієнта.

Крок 3
У цьому прикладі нуклеаза деградує одну нитку донорського фрагмента, а решта нитка ДНК потрапляє до реципієнта. Специфічні для компетенції однониткові ДНК-зв'язуючі білки зв'язуються з ниткою донорської ДНК, щоб запобігти її деградації в цитоплазмі.

Крок 4
Білки ReCa сприяють генетичному обміну між фрагментом ДНК донора та ДНК реципієнта. Це передбачає розрив і возз'єднання парних сегментів ДНК.
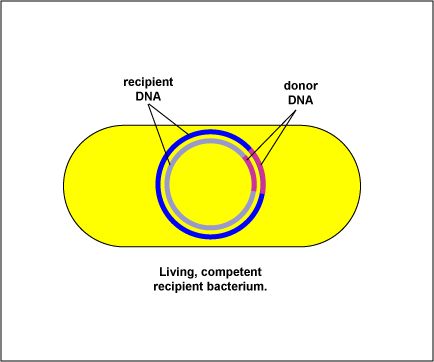
Крок 5
Трансформація завершена.

Мутації
Мутації (лат. mutatio — зміна) — це стійкі дискретні (переривчасті) зміни генетичного апарату, що виникають раптово і впливають на ознаки й властивості організмів та вірусів; успадковуються поколіннями клітин або нащадками. Термін запровадив Г. Де Фрізу в 1901 р.

Види
Мутації можна класифікувати за характером прояву, місцем виникнення, за характером змін спадкового апарату, за походженням тощо. За характером прояву мутації поділяють на домінантні та рецесивні. Більшість з них є рецесивними і не проявляються у гетерозигот. Домінантні мутації — це мутації, що проявляються у гомо- і гетерозиготного організму та з'являються вже в першому поколінні. Наприклад, полідактилія, або багатопалість, є мутацією, що успадковується за аутосомно-домінантним типом. Рецесивні мутації — це мутації, що не проявляються у гетерозигот і з'являються тільки в другому і наступних поколіннях. Так, у людини рецесивний характер має така мутація, як альбінізм. За впливом на життєдіяльність розрізняють летальні, сублетальні та нейтральні мутації. Летальні мутації спричиняють загибель організмів ще до моменту народження або до настання здатності до розмноження. У людини до таких мутацій належить гемофілія. Мутації, які знижують життєздатність особин і частково або повністю зупиняють розвиток, називаються сублетальними. Якщо ж мутації за звичних умов не впливають на життєздатність організмів, то їх називають нейтральними. За місцем виникнення мутації класифікують на соматичні та генеративні. Соматичні мутації виникають у соматичних диплоїдних клітинах, часто не передаються спадково і обмежені тією особиною, в якої виникли. Наприклад, у людини соматичними мутаціями є мозаїцизм або пухлинне переродження клітин. Мутації, що виникли в гаметах чи клітинах, з яких вони утворюються, і передаються нащадкам у випадку їхньої участі в заплідненні, називають генеративними мутаціями. За характером змін спадкового апарату мутації бувають генними, хромосомними та геномними. Генні мутації — стійкі зміни окремих генів, спричинені порушенням послідовності нуклеотидів у молекулах нуклеїнових кислот. Ці мутації виникають внаслідок випадання певних нуклеотидів, появи зайвих, зміни порядку їх розташування. Генними мутаціями в людини є полідактилія, альбінізм. Хромосомні мутації — це мутації, що виникають у результаті перебудови хромосом. Вони можуть виникати як в межах однієї хромосоми, так і між гомологічними та негомологічними хромосомами. Прикладом хромосомних мутацій є синдром «котячого крику» (46,5—), гострий лейкоз (46,21—). Геномні мутації — це мутації, що пов'язані зі зміною кількості наборів хромосом. Основними видами геномних мутацій є: збільшення кількості хромосомних наборів (поліпроїдія), зменшення кількості хромосомних наборів, зміна числа хромосом окремих пар (анеуплоїдія). Прикладами геномних мутацій є синдром Дауна (47,21+) в людини.

Точкові
Точкові Мутації — результат зміни молекулярної структури ДНК чи рибонуклеїнової кислоти (РНК) або цитологічно непомітних хромосомних мікроаберацій, що легко виявляються за допомогою біохімічного чи гібридологічного аналізу.
a
Міссенс-мутації
Міссенс-мутація (англ. Missense mutation) — це точкова мутація, заміна нуклеотиду призводить до утворення кодону, який кодує іншу амінокислоту. Таким чином відбувається заміна однієї амінокислоти на іншу. Ці мутації можуть мати різні наслідки для структури та функції білка, що кодується.

Зміни структури ДНК
Зміни структури ДНК можуть включати мутації, які впливають на зв'язок між нуклеотидами або їхню послідовність. Це може призвести до змін у структурі білків, оскільки змінюється код для синтезу амінокислот.
Зміни функціональності білка
Заміна амінокислоти може призвести до зміни третинної або четвертинної структури білка, що, в свою чергу, вплине на його функцію.
aНеправильне згортання білка
Заміна амінокислоти може порушити нормальний процес згортання білка, що призводить до утворення неправильно згортаних форм білка. Це може впливати на його функцію та стабільність.
a
Нейродегенеративні захворювання
Нейродегенеративні захворювання - група патологій нервової системи, яка включає хвороби, пов'язані з ураженням і загибеллю нейронів. До цих станів відносять хворобу Альцгеймера, Паркінсона, розсіяний склероз і багато інших. Нейродегенеративні захворювання поступово призводять до інвалідизації пацієнта.
Руйнування нейронів
Міссенс мутації можуть призводити до руйнування нейронів, оскільки аномальні білки не здатні виконувати свої функції, що впливає на нервову систему і може викликати серйозні нейропатології.

Погіршення пам'яті
Погіршення пам'яті є наслідком нейродегенеративних захворювань, викликаних міссенс мутаціями. Втрата функціональності нейронів та зниження пластичності синапсів призводить до труднощів у запам'ятовуванні та відтворенні інформації.
Втрата самостійності хворого
Хворі на нейродегенеративні захворювання, такі як хвороба Гантінгтона, можуть втратити здатність до самообслуговування через порушення моторики та когнітивних функцій.
Прогресування хвороби
Прогресування хвороби відбувається через накопичення ушкоджень у нейронах та інші патологічні зміни, спричинені міссенс мутаціями. Це може призводити до більш тяжких форм захворювання та необхідності в медичному догляді.
Пріонні захворювання
Пріонні захворювання — це група нейродегенеративних хвороб людини та тварин, до яких відносяться хвороба Крейтцфельдта—Якоба (ХКЯ), хвороба Герстманна—Штраусслера—Шейнкера (ХГШШ) у людей, та губчаста енцефалопатія великої рогатої худоби (ГЕВРХ, або «коров'ячий сказ») та скрепі у овець.
a
Хвороба Гантінгтона
Хворо́ба Га́нтінгтона — генетичне захворювання нервової системи, яке уражає людину у віці 35-50 років внаслідок змін у гені IT-15, який знаходиться на 4-ій хромосомі. Призводить до атрофії стріатума, а на пізній стадії — до атрофії кори головного мозку.
aЗахворюваня через дисфункціональні білки
Міссенс мутації можуть викликати дисфункцію білків, що, у свою чергу, може призводити до розвитку різних захворювань. Наприклад, зміни в структурі білка, відповідального за метаболізм, можуть призвести до метаболічних порушень.
a
Втрата функціональності білка
Міссенс мутації може призвести до втрати функціональності білка, якщо нова амінокислота не виконує необхідних функцій або порушує структуру активного центру білка.
Нові властивостей білка
У деяких випадках, міссенс мутації може надавати білку нові функції або властивості. Це може бути корисним, якщо новий білок краще адаптується до умов середовища або має нові біохімічні властивості.
Нонсенс-мутації
Нонсенс-мутація — точкова мутація в послідовності ДНК, яка призводить до появи стоп-кодону (нонсенс-кодону) в транскрибованій мРНК. Результатом є неповний і зазвичай нефункціональний продукт цієї мРНК.

Синтетичні мутації
Синтетичні мутації — це мутації, які спеціально створені або виведені в лабораторних умовах шляхом модифікації ДНК. Вони можуть бути отримані за допомогою різних методів, таких як:Генетичне редагування (наприклад, CRISPR-Cas9), що дозволяє точно вносити зміни в геном.Хімічні мутагени, які можуть змінювати структуру ДНК.Інші техніки молекулярної біології, що використовуються для створення певних мутацій з метою вивчення функцій генів або виведення нових ознак у організмах.Синтетичні мутації використовуються в дослідженнях для кращого розуміння генетичних механізмів, а також в агрономії, медицині та біотехнології.
a
Делеції
Делеція (делеція гена) в генетиці — структурна мутація (хромосомна аберація), при якій вилучається частина хромосоми або послідовність ДНК. При делеції виникає втрата генетичного матеріалу. При цьому може бути вилучена будь-яке число нуклеотидів, від одної основи до великого шматка хромосоми.
Інсерції
Інсерції у мутаціях — це тип генетичної мутації, при якому до ДНК додається один або кілька нуклеотидів, що може змінити структуру гена та вплинути на синтез білків. Інсерції можуть порушувати нормальні функції генів або викликати зсув рамки зчитування.
Хромосомні перебудови
Абера́ції хромосо́м (від лат. aberratio — відхилення, віддалення), мутації хромосом, хромосомні перебудови — порушення структури (структурна перебудова) хромосом (хроматид), що супроводжується їхнім розривом, після якого зазвичай розірвані кінці з'єднуються у нових поєднаннях (комбінаціях).

Технології
5 липня 1996 року в Шотландії народилася перша успішно клонована з дорослої клітини тварина — вівця з ідентифікаційним кодом 6LL3. Її назвали Доллі на честь кантрі-співачки Доллі Партон.Біолог Ян Вільмут, очільник групи дослідників Рослинського інституту, оголосив про клонування вівці тільки через пів року після того, як це сталося — 22 лютого 1997-го. Вчені не квапилися з оголошенням, щоб воно збіглося в часі з виходом статті в журналі Nature. У 21-му ст. відбувся ряд відкриттів в області генетики.
NGS
NGS, або покоління секвенування, – це група сучасних технологій, які дозволяють швидко секвенувати ДНК та РНК. На відміну від традиційних методів секвенування, таких як Sanger-секвенування, NGS дозволяє отримувати величезну кількість даних за короткий час і за нижчою ціною.
a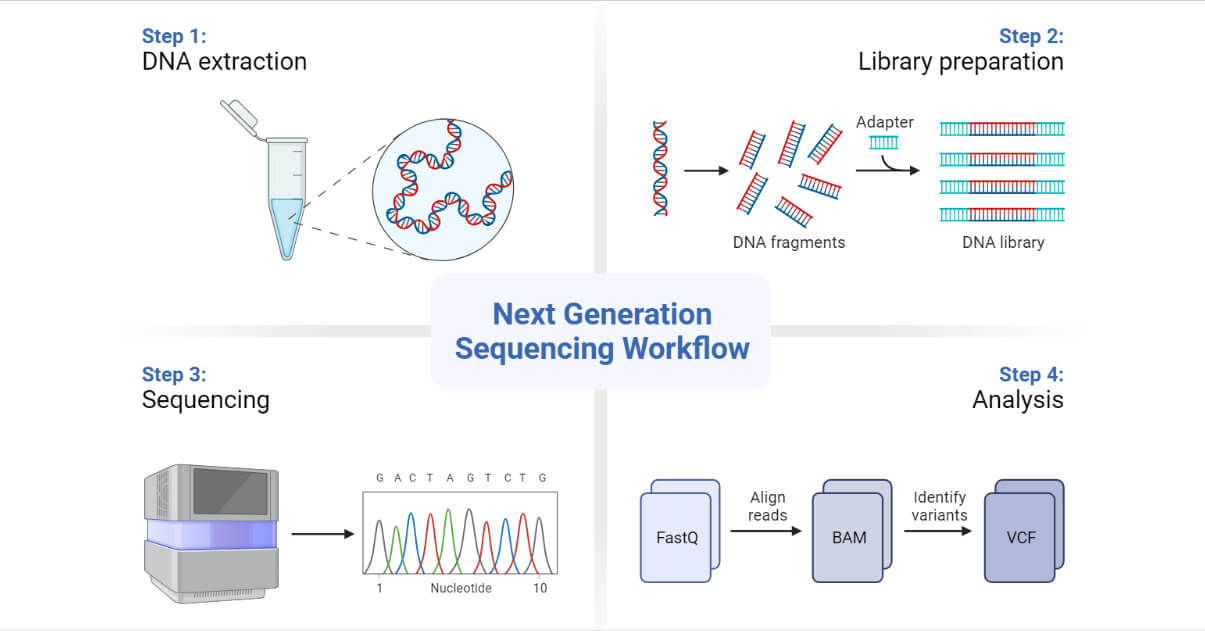
Принцип роботи
Принцип роботи здійснюється в чотири етапи.
Етап 1
Спочатку зразок ДНК або РНК фрагментується, а потім до фрагментів додаються адаптери – короткі послідовності ДНК, які дозволяють зв'язуватися з платформою для секвенування.
Етап 2
Отримані фрагменти ДНК ампліфікуються методом ПЛР (полімеразна ланцюгова реакція) для отримання достатньої кількості матеріалу.
Етап 3
Після підготовки бібліотеки фрагменти ДНК секвенуються за допомогою платформи NGS. Технології секвенування можуть включати секвенування за синтезом (Illumina), секвенування за методом розпізнавання (Ion Torrent) або секвенування на основі нанопор (Oxford Nanopore).

Етап 4
Після секвенування отримані дані обробляються та аналізуються для отримання інформації про геном, експресію генів, варіації, поліморфізми, та інші генетичні особливості.
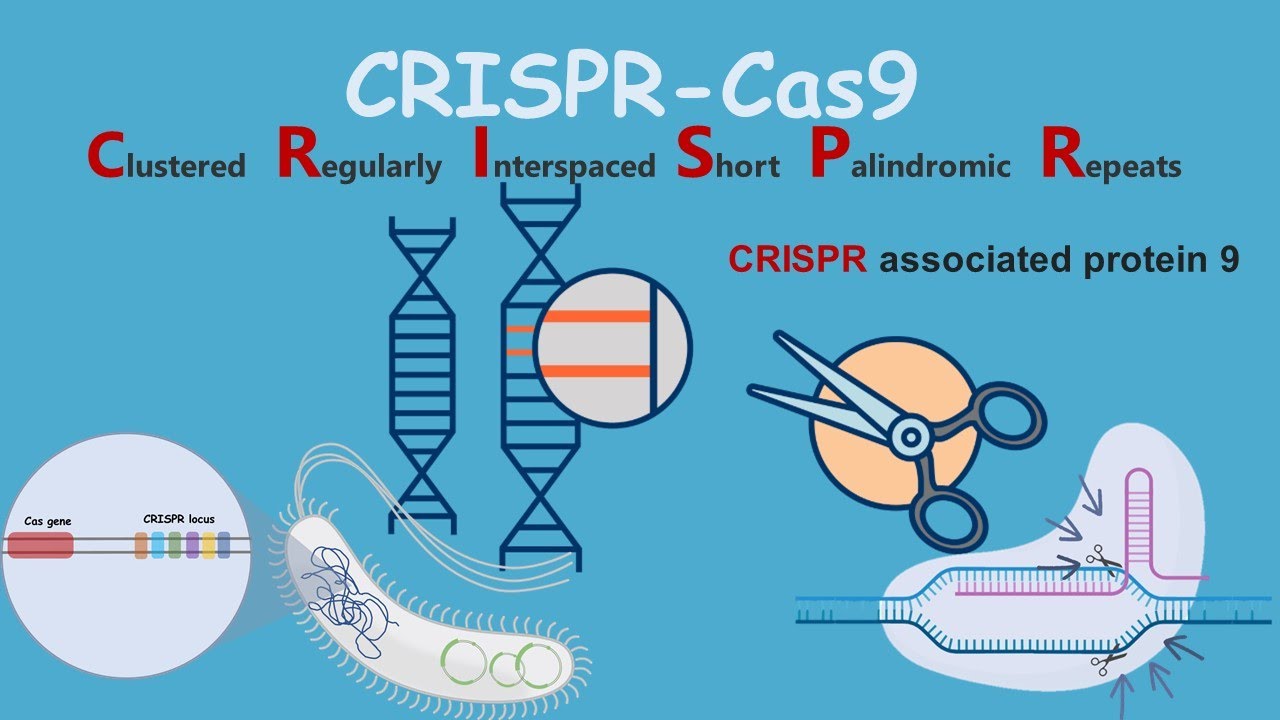
CRISPR-Cas9
CRISPR-Cas9 – це революційна технологія редагування генів, яка дозволяє точно модифікувати ДНК у живих організмах. Вона базується на природній системі захисту, знайденій у бактерій, що захищають їх від вірусів.

Принцип роботи
Принцип роботи здійснюється в п'ять етапів.
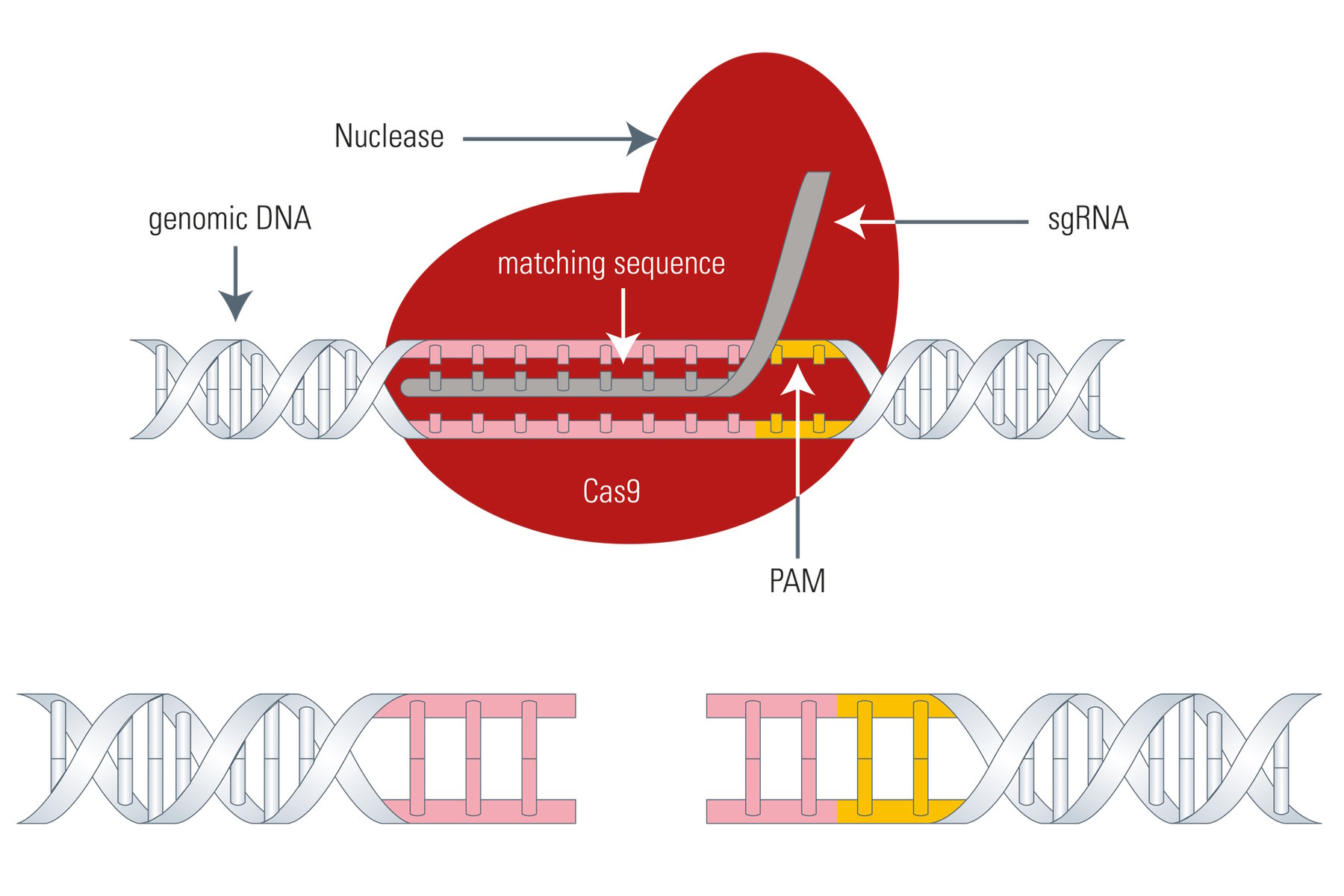
Вибір цільового гена
Спочатку розробляється коротка РНК (sgRNA), яка комплементарна до цільової послідовності ДНК у геномі. Ця РНК служить провідником для Cas9.
Утворення комплексу
Утворення комплексу: sgRNA зв'язується з білком Cas9, утворюючи комплекс.
Вступ до клітини
Комплекс CRISPR-Cas9 вводиться в клітину, де sgRNA зв'язується з цільовою ДНК.

Розщеплення ДНК
Cas9 здійснює розщеплення дволанцюгової ДНК у вказаному місці, викликаючи утворення «двох розривів».

Виправлення або модифікація
Після розщеплення ДНК клітина намагається відновити цілісність молекули ДНК. Цей процес можна використати для введення нових генетичних матеріалів (наприклад, векторів) або для редагування вже наявних генів шляхом включення нових послідовностей ДНК.