Sirve de reserva energética y se encuentra en dos tejidos
Glucógeno
GLUCOGENOGÉNESIS
GLUCOSA
G – 6 – P
G – 1 – P
URIDINA DIFOSFATO GLUCOSA
ENLACES α 1,4
ENLACES α 1,6
GLUCÓGENO
GLUCOGENÓLISIS
GLUCÓGENO
G – 1 – P
G – 6 – P
GLUCOSA
ENLACES α 1,6
GLUCOSA
Polímero de cadenas ramificadas de glucosa.
Hígado
Sangre
Glucosa
Glucosa en Sangre
Glucosa
Glicolisis
Piruvato
Alanina
Proteina Muscular
Aminoacidos
NH4+
Glutamato
alfa Cetoglutarato
Alanina en Sangre
Alfa Cetoglutarato
Piruvato
Glutamato
NH4+
Urea
Fermentacion láctica
Lactato
Lactato en sangre
Lactato
Glucógeno
Oxaloacetato
Malato
Malato
Oxaloacetato
FOSFO – ENOL – PIRUVATO
2 – FOSFO – GLICERATO
3 – FOSFO – GLICERATO
1,3 DIFOSFOGLICERATO
GLICERALDEHIDO – 3 – P
FRUCTOSA – 1,6 – BIP
FRUCTOSA – 6 – P
GLUCOSA – 6 – P
Hígado
Músculo
Músculo
HÍGADO
Leonardo Bortoluzzi
Vasti Pelayo
Hiago Tavares
Integrantes
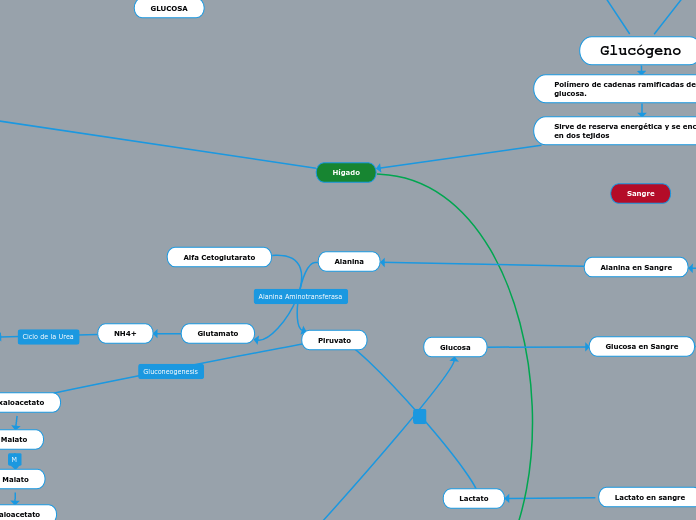
A diferencia de la glucogenólisis, que puede responder rápidamente al estímulo hormonal, la gluconeogénesis es una respuesta más lenta que depende de cambios en la expresión génica y que alcanza una actividad máxima en un período de horas. La gluconeogénesis requiere una fuente de energía para la biosíntesis y una fuente de carbonos para la
formación del esqueleto de la molécula de glucosa. La energía es proporcionada por el metabolismo de los ácidos grasos
liberados por el tejido adiposo. La estructura carbonada es suministrada por tres fuentes principales:
El lactato producido en tejidos como los hematíes y el músculo.
Los aminoácidos derivados de las proteínas musculares.
El glicerol liberado a partir de los triglicéridos durante la lipólisis en el tejido adiposo.
Entre éstos, la proteína muscular es el p re cursor más importante de la glucosa sanguínea durante el ayuno y la inanición,
ya que la velocidad de la gluconeogénesis está limitada a menudo por la disponibilidad del sustrato, incluida la tasa de
proteólisis en el músculo o, en algunos casos, la masa muscular.
La vía de la glucogenólisis comienza con la eliminación de los abundantes residuos externos de glucosa unidos mediante enlaces α 1 —>4 en el glucógeno. Esto se realiza por la glucógeno fosforilasa, una enzima que utiliza el fosfato citosólico y libera glucosa del glucógeno en forma de Glc-1-P. La Glc-l-P se isomeriza por la
fosfoglucomutasa a Glc-6-P, situándola al inicio de la vía glucolítica.
En el hígado, la glucosa se libera a partir de la Glc-6-P por la glucosa-6-fosfatasa, y la glucosa se incorpora a la
sangre mediante el transportador GLUT-2.
La fosforilasa es específica para los enlaces glucosídicos α 1—>4; no puede escindir los enlaces α 1 —>6.
La enzima desramificante rompe los enlaces α 1 —>6 hasta llegar a glucosa.
Glucogenesis Hepatica
Conversión de Glc-6-P en glucosa-1-fosfato (Glc-l-P) por la fosfoglucomutasa.
Activación de la Glc-l-P para formar el azúcar nucleotídico, uridina difosfato glucosa (UDP)-glucosa mediante la enzima UDP-glucosa pirofosforilasa.
Transferencia de glucosa desde la UDP-Glc al glucógeno en un enlace al—>4 mediante la glucógeno sintasa, un miembro de la clase de enzimas conocida como glucosil transferasas.
Cuando la longitud de la cadena a 1 —>4 es mayor de 8 residuos, la enzima ramificante del glucógeno, una transglucosilasa, transfiere parte de los azúcares de los enlaces a 1—>4 a una rama al—>6, estableciendo el estadio para el alargamiento continuo de ambas cadenas en a 1—>4 hasta que, a su vez, se hacen lo suficientemente largas para la transferencia por la enzima ramificante.