

Simbiosis
Ninguna especie vive aislada, pero a menudo la asociación con otras especies es especialmente estrecha: para muchos organismos, el hábitat que ocupan es un individuo de otra especie. Los parásitos viven dentro de las cavidades corporales o incluso en las células de sus huéspedes, las bacterias fijadoras de nitrógeno viven en nódulos en las raíces de las plantas leguminosas, y así sucesivamente. Simbiosis ("vivir juntos") es el término que se ha acuñado para esas asociaciones físicas tan cercanas entre especies, en las que un "simbionte" ocupa un hábitat proporcionado por un "huésped". Sin embargo, los parásitos generalmente se excluyen de la categoría de simbiontes, que se reserva en su lugar para las interacciones donde hay al menos la sugerencia de mutualismo.
Protectores mutualistas

Agricultura o ganadería
Al menos en términos de extensión geográfica, algunos de los mutualismos más dramáticos son los de la agricultura humana. El número de plantas individuales de trigo, cebada, avena, maíz y arroz, y las áreas que ocupan estos cultivos, superan ampliamente lo que habría estado presente si no se hubieran cultivado. El aumento de la población humana desde la época de los cazadores-recolectores es una medida de la ventaja recíproca del Homo sapiens. Incluso sin hacer el experimento, podemos imaginar fácilmente el efecto que la extinción de los humanos tendría en la población mundial de plantas de arroz o el efecto de la extinción de las plantas de arroz en la población de humanos. Comentarios similares se aplican a la domesticación de bovinos, ovinos y otros mamíferos.

Dispersión de semillas y polen
Muchas especies de plantas usan animales para dispersar sus semillas y polen. Alrededor del 10% de todas las plantas con flores poseen semillas o frutos que llevan anzuelos, púas o pegamentos que se adhieren a los pelos, cerdas o plumas de cualquier animal que entre en contacto con ellos. Con frecuencia son una irritación para el animal, que a menudo se limpia y elimina si puede, pero generalmente después de llevarlos a cierta distancia. En estos casos, el beneficio es para la planta (que ha invertido recursos en mecanismos de pegamento) y no hay recompensa para el animal.
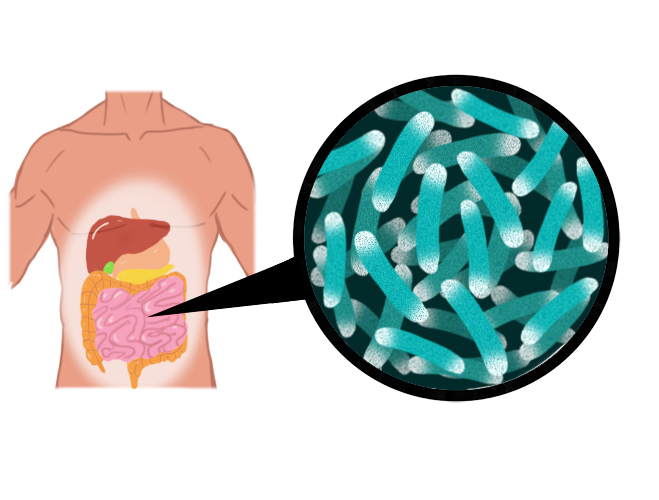
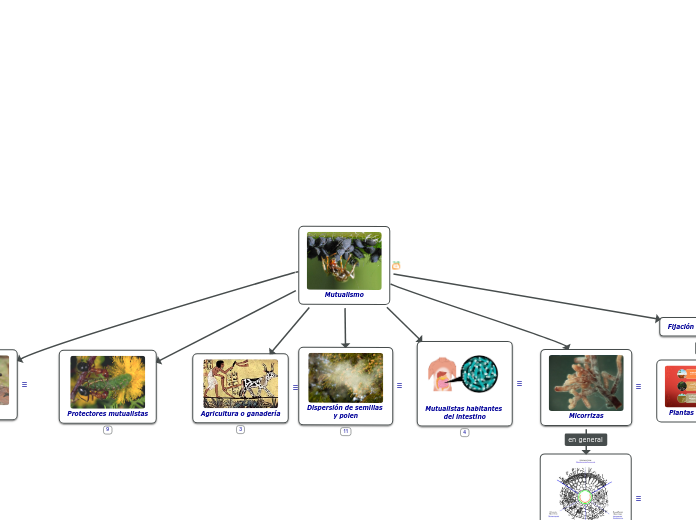
Mutualistas habitantes del intestino
La mayoría de los mutualismos discutidos hasta ahora han dependido de patrones de comportamiento, donde ninguna de las especies vive completamente "dentro" de su pareja. En muchos otros mutualismos, uno de los socios es un eucariota unicelular o bacteria que se integra más o menos permanentemente en la cavidad corporal o incluso en las células de su compañero multicelular. La microbiota que ocupa partes de los canales alimenticios de varios animales son los simbiontes extracelulares más conocidos.
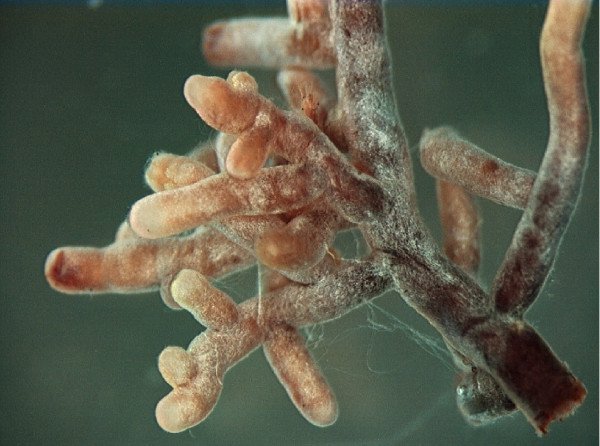
Micorrizas
La mayoría de las plantas superiores no tienen raíces, tienen micorrizas, mutualismos íntimos entre los hongos y el tejido radicular. Las plantas de solo unas pocas familias, como las Cruciferae, son excepciones. En términos generales, las redes fúngicas en micorrizas capturan nutrientes del suelo, que transportan a las plantas a cambio de carbono. Muchas especies de plantas pueden vivir sin sus hongos micorrícicos en suelos donde ni los nutrientes ni el agua son limitantes, pero en el duro mundo de las comunidades de plantas naturales, las simbiosis, si no son estrictamente obligatorias, son 'ecológicamente obligatorias': es decir, necesarias si los individuos deben sobrevivir en la naturaleza (Buscot et al., 2000).

3 Tipos
En general, se reconocen tres tipos principales de micorrizas. Las micorrizas arbusculares se encuentran en aproximadamente dos tercios de todas las especies de plantas, incluidas la mayoría de las especies no leñosas y los árboles tropicales. Los hongos ectomicorrícicos forman simbiosis con muchos árboles y arbustos, dominando los bosques boreales y templados y también algunas selvas tropicales. Finalmente, las micorrizas ericoides se encuentran en las especies de plantas dominantes de brezales.

Ectomicorrizas
En las ectomicorrizas (ECM), las raíces infectadas generalmente se concentran en la capa de arena del suelo. Los hongos forman una vaina de grosor variable alrededor de las raíces. A partir de ahí, las hifas irradian hacia la capa de arena, extrayendo nutrientes y agua y también produciendo grandes cuerpos fructíferos que liberan enormes cantidades de esporas transportadas por el viento. El micelio fúngico también se extiende hacia adentro desde la vaina, penetrando entre las células de la corteza de la raíz para dar un contacto íntimo de célula a célula con el huésped y estableciendo una interfaz con una gran área de superficie para el intercambio de los productos de la fotosíntesis, agua del suelo y nutrientes entre la planta huésped y su compañero fúngico.

Microrrizas arbusculares
Las micorrizas arbusculares (AM) no forman una vaina sino que penetran en las raíces del huésped. Las raíces se infectan por el micelio presente en el suelo o por los tubos de infección que se desarrollan a partir de esporas asexuales, que son muy grandes y se producen en pequeñas cantidades: un contraste sorprendente con los hongos ECM. Inicialmente, el hongo crece entre las células huésped, pero luego entra en ellas y forma un "arbúsculo" intracelular finamente ramificado.
Absorción de fósforo y otras ventajas
Ha habido una tendencia a enfatizar la facilitación de la absorción de fósforo como el principal beneficio para las plantas de las simbiosis MA (el fósforo es un elemento altamente inmóvil en el suelo y, por lo tanto, con frecuencia limita el crecimiento de las plantas), pero la verdad parece ser más complejo que esto, con beneficios demostrados también en la absorción de nitrógeno, la protección de patógenos y herbívoros y la resistencia a metales tóxicos (Newsham et al., 1995). Ciertamente, hay casos en los que la entrada de fósforo está fuertemente relacionada con el grado de colonización de las raíces por MA.
Campanilla azul
Esto se ha demostrado para la campanilla azul, Hyacinthoides non-scripta, a medida que avanza la colonización durante su fase de crecimiento subterráneo desde agosto a febrero hasta su fase fotosintética sobre el suelo (Figura 8.14a). De hecho, las campanillas azules cultivadas sin MA no pueden absorber el fósforo a través de su sistema de raíces poco ramificado (Merryweather y Fitter, 1995).Curvas ajustadas a las tasas de entrada de fósforo (línea discontinua, eje izquierdo) y colonización de raíces por micorrizas arbusculares (línea continua, eje derecho) en la campanilla azul, Hyacinthoides non-scripta, durante una sola temporada de crecimiento. La absorción de fósforo parece estar fuertemente vinculada a la colonización de raíces por los hongos.

Hyacinthoides non-scripta
Protección contra patógenos
Por otro lado, un conjunto de experimentos examinó el crecimiento anual hierba Vulpia ciliata ssp. ambigua (Figura 8.14b) en la que se cultivaron plántulas de Vulpia con un MA (Glomus sp.), con el hongo patógeno Fusarium oxysporum, con ambos y con ninguno. El crecimiento no se mejoró solo con Glomus, pero Fusarium perjudicó el crecimiento en ausencia de Glomus. Cuando ambos estuvieron presentes, el crecimiento volvió a los niveles normales. Claramente, la micorriza no benefició la economía de fósforo de la Vulpia, pero sí la protegió de los efectos nocivos del patógeno.
Protección contra patógenos
Los efectos de una combinación factorial de Fusarium oxysporum (Fus, un hongo patógeno) y un hongo AM, Glomus sp. (Glm) en el crecimiento (longitud de la raíz) de las plantas Vulpia. Los valores son medias de 16 repeticiones por tratamiento; las barras son errores estándar; el asterisco significa una diferencia significativa en P <0.05 en una comparación por pares de Fisher. En este caso, el beneficio proporcionado por los hongos AM no parece ser una mejora en la absorción de nutrientes sino una protección contra el patógeno.

Vulpia
Fusarium oxysporum

Glomus
Especie
La diferencia clave parece ser que Vulpia, a diferencia de la campanilla azul, tiene un sistema de raíces altamente ramificado (Newsham et al., 1995). Las plantas con raíces finamente ramificadas tienen poca necesidad de captura adicional de fósforo, pero el desarrollo de esa misma arquitectura de raíces proporciona múltiples puntos de entrada para los patógenos de las plantas. En tales casos, es probable que las simbiosis MA hayan evolucionado con énfasis en la protección de las plantas. Por el contrario, los sistemas de raíces con pocos meristemos laterales y de crecimiento activo son relativamente invulnerables al ataque de patógenos, pero estos sistemas de raíces son pobres en la búsqueda de fósforo. Aquí, es probable que las simbiosis MA hayan evolucionado con énfasis en la captura de fósforo.
Fijación de Nitrógeno
Plantas mutualistas